This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
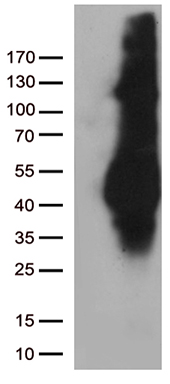
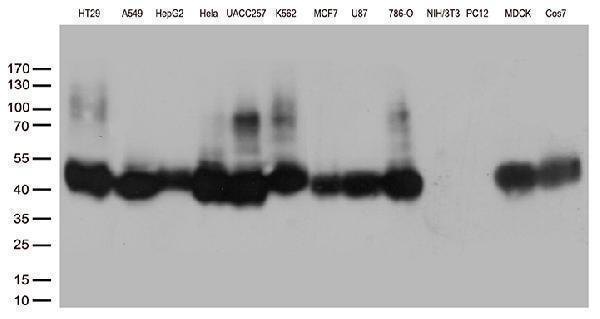

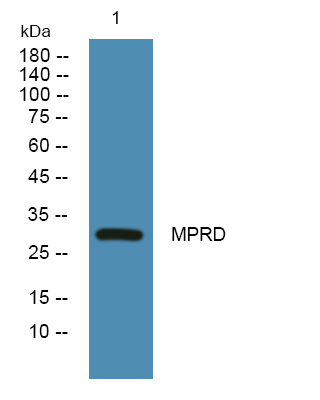
Facts about Cation-dependent mannose-6-phosphate receptor.

.
| Human | |
|---|---|
| Gene Name: | M6PR |
| Uniprot: | P20645 |
| Entrez: | 4074 |

| Belongs to: |
|---|
| No superfamily |

000 Man6PR; 46 kDa mannose 6-phosphate receptor; 46-kDa mannose 6-phosphate receptor; cation-dependent mannose-6-phosphate receptor; CD Man-6-P receptor; CDM6PR; CD-M6PR; CD-MPR; FLJ32994; M6PR; mannose-6-phosphate receptor (cation dependent); MPR 46; MPR46; MPRD; Mr 46; SMPR
Mass (kDA):
30.993 kDA

| Human | |
|---|---|
| Location: | 12p13.31 |
| Sequence: | 12; NC_000012.12 (8940361..8949761, complement) |
Lysosome membrane; Single-pass type I membrane protein.



PMID: 2441386 by Pohlmann R., et al. Cloning of a cDNA encoding the human cation-dependent mannose 6- phosphate-specific receptor.
PMID: 1849818 by Klier H.J., et al. Isolation and analysis of the human 46-kDa mannose 6-phosphate receptor gene.