This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
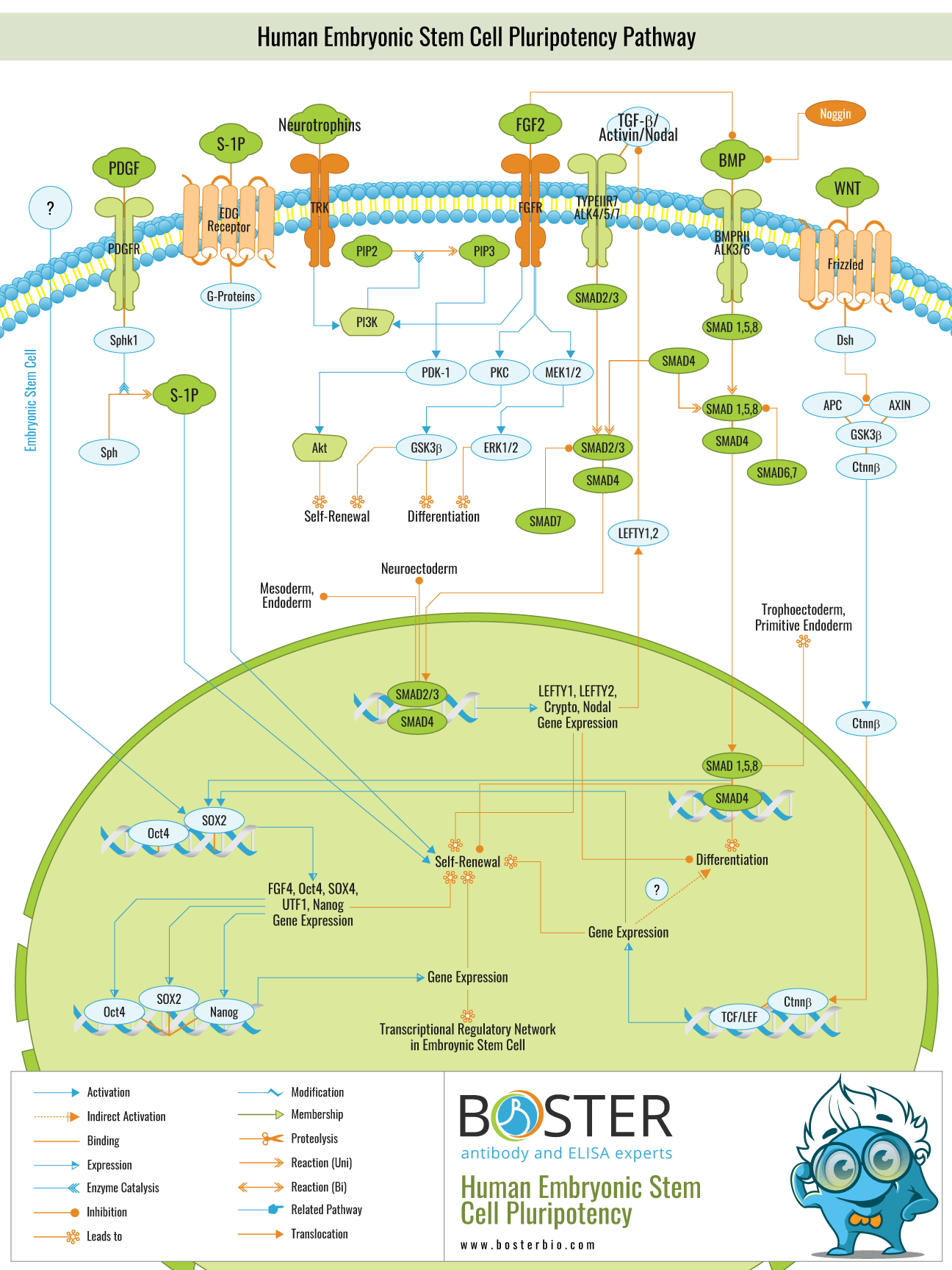
Through a variety of molecular signaling pathways, human embryonic stem cells (ESCs) maintain their pluripotency. Extracellular proteins such as PDGF, Neurotrophins, FGF, BMPs, and WNT control signaling cascades involved in stem cell renewal and differentiation. BMP signaling, for example, activates SMAD signaling pathways. Once SMAD transcription factors are translocated to the nucleus, they regulate self-renewal genes such as LEFTY1, Cripto, and Nodal. Other transcription factors such as Oct4, Soc2, and Nanog are also expressed as part of transcriptional regulatory networks that contribute to the maintenance of pluripotency or differentiation.
Other transcription factors such as Oct4, Soc2, and Nanog are also expressed as part of transcriptional regulatory networks that contribute to the maintenance of pluripotency or differentiation.
Human pluripotent stem cells derived from embryonic cells and those derived from reprogrammed somatic cells share a number of characteristics, including the ability to proliferate indefinitely and to differentiate into a wide variety of cell types. However, it remains unknown whether both cell types rely on similar mechanisms to maintain pluripotency and regulate differentiation. Any differences in these mechanisms would imply that reprogramming fibroblasts to generate induced pluripotent stem cells (iPSCs) results in novel pluripotent states. Current methods for expanding and differentiating human embryonic stem cells (ESCs) may not be directly applicable to human iPSCs in that case. However, we demonstrate here that human iPSCs rely on activin/nodal signaling to maintain pluripotency and thus are mechanistically similar to human ESCs. Additionally, we demonstrate that the growth factors required and sufficient for specification of human embryonic stem cells into extraembryonic tissues, neuroectoderm, and mesendoderm also drive differentiation of human iPSCs into the same tissues.
Notably, experiments were conducted in a completely chemically defined medium that was free of factors that could obscure the analysis of developmental mechanisms or render the resulting tissues unsuitable for future clinical applications. Together, these findings demonstrate that human iPSCs and human ESCs both rely on mechanisms similar to those found in human ESCs to maintain pluripotency and control differentiation, demonstrating that these two pluripotent cell types are functionally equivalent.
As with embryos, differentiation in hESCs is triggered by extracellular developmental cues such as morphogens or cytokines. These morphogens or cytokines interact with their cognate plasma membrane-bound receptors and activate specific intracellular signaling pathways. Signaling pathways are activated through a series of phosphorylation events that ultimately result in the regulation of specific transcription factors. These transcription factors, in turn, can recruit additional cofactors and directly regulate downstream gene transcription. Additionally, transcription factors can recruit histone modifying and chromatin remodeling enzymes to reorganize the epigenetic structure, rendering pluripotency genes inaccessible for transcription and repressed, while lineage-specific genes become accessible for transcription and activated. This series of events culminates in the expression of lineage-specific proteins such as transcription factors and structural proteins, resulting in a change in the cell's morphology. Additionally, pluripotency-associated transcription factors and other pluripotency-associated genes are repressed indefinitely, completing the differentiation process. Thus, differentiation is a fairly complex series of events that are regulated by signaling pathways, transcription factors, epigenetic factors, and lineage-specific proteins.
While significant knowledge about each of these functional groups has been accumulated in isolation (i.e., signaling pathways, transcription factors, epigenetic factors, and lineage-specific proteins), very little is known about their interactions, particularly in the context of hESC differentiation. In part, interactions between these groups confer lineage specificity on the differentiation process and mediate the development of specific tissues in response to morphogen exposure of hESCs.