This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
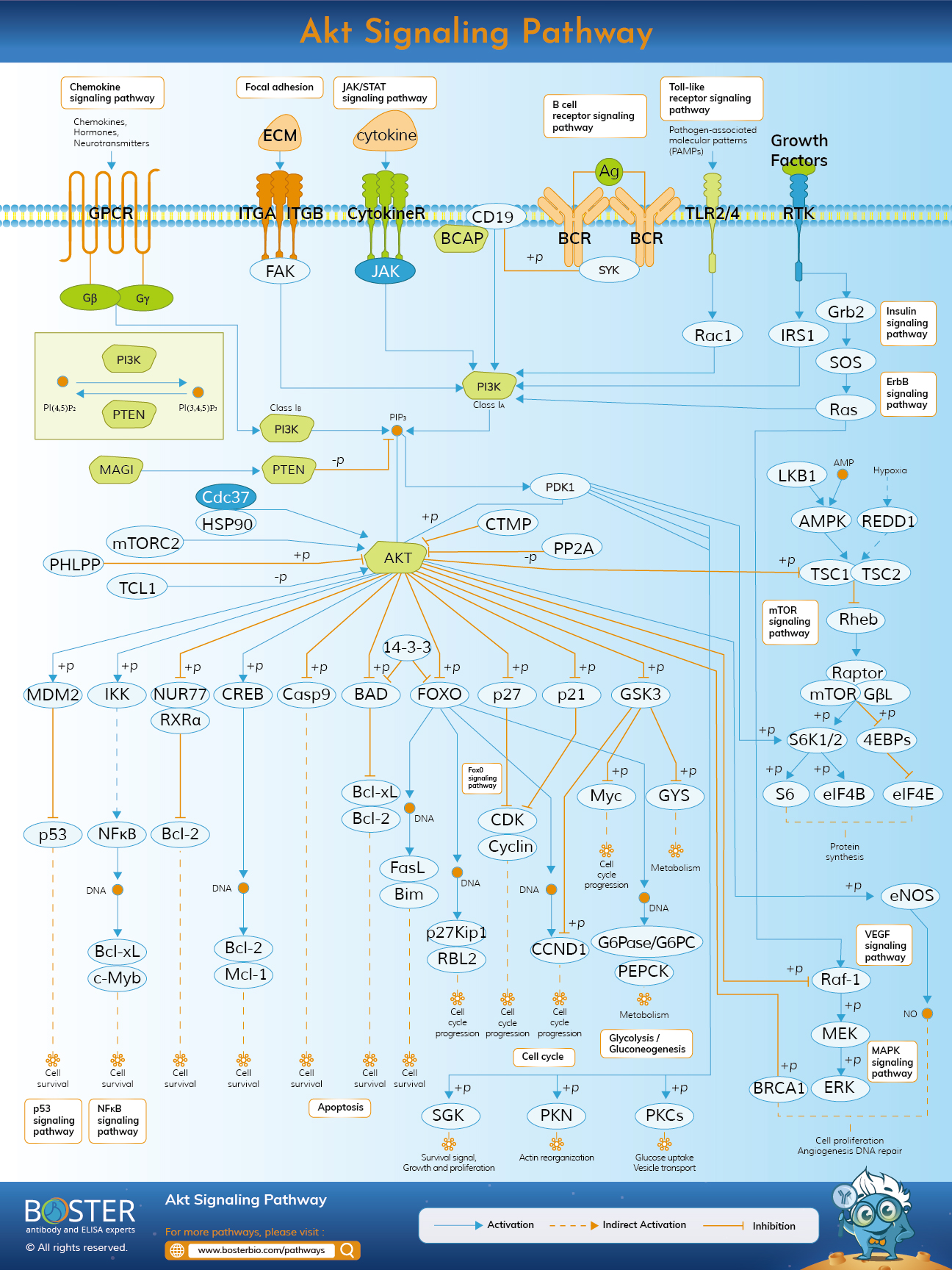
Akt signaling has an important role in the regulation of signal transduction and biological processes such as cell proliferation, apoptosis, metabolism and angiogenesis.
The origin of the PKB / Akt research dates back to the discovery by Staal and his colleagues in 1977 that it is an oncogene in a previously undescribed virus called ATK8. And this cell-derived cancer sequence was isolated and named akt. In 1991, three independent research teams identified the gene for PKB / Akt. These three cloning articles established PKB / Akt as a new type of widely expressed phosphoprotein kinase and paved the way for future research on the role of PKB / Akt in different cellular processes. Cantley's group isolated an enzyme called phosphatidylinositol 3 kinase (PI3K) in 1990. In 1995, Richard Roth and his colleagues reported that insulin activated Akt. Later, some studies have shown that the membrane phospholipids produced by PI3K are a complete element required to activate PKB/Akt.
Akt pathway is an intracellular signal transduction pathway that can promote metabolism, proliferation, cell survival, growth, and angiogenesis in response to extracellular signals. This is mediated by serine and / or threonine phosphorylation of a number of downstream substrates. The key proteins involved are phosphatidylinositol 3 kinase (PI3K) and Akt / protein kinase B.
AKT, also known as protein kinase B, is a specific serine/threonine protein kinase that plays a key role in multiple cellular processes. Once activated, Akt regulates functions by activating or inhibiting the phosphorylation of various proteins involved in cell growth, proliferation, motility, adhesion, new blood vessel formation, and cell death.
Akt / PKB is characterized by its similarity to protein kinases A (PKA) and C (PKC) and the retroviral oncoprotein virus akt (vakt). Structurally, Akt contains three domains: amino-terminal (N-terminal), central, and carboxy-terminal (C-terminal) fragments. The N-terminal domain is the homology domain (PH) of the Plekster protein, composed of 100 amino acids, similar to other domains in 3 phospholipid binding molecules, and can interact with membrane lipid products such as phosphatidylinositol (3,4, 5) triphosphate. (PIP3) and phosphatidylinositol 4,5 diphosphate (PIP2). The kinase domain is very similar to the AGC protein kinase that shares the regulatory threonine residue Thr308. Phosphorylation of this residue activates Akt. The C-terminal groove is composed of 40 amino acids, forming a hydrophobic region that contains the regulatory serine residue Ser473.
There are three highly related Akt subtypes (Akt1, Akt2, and Akt3) that phosphorylate substrates containing the common phosphorylation motif of RxRxxS/T. Akt isotypes share many substrates, but isotype-specific Akt substrates have also been identified. For example, all Akt subtypes can phosphorylate PRAS40 (40 kDa proline-rich Akt substrate), but only Akt1 can phosphorylate the actin-related protein palladin.
This includes Akt activation, PI3K activation, and Phosphoinositide formation.
Phosphatidylinositol 3 kinase (PI3K) can be divided into three subtypes based on its structure and substrate specificity: Class I, Class II, and Class III. Among these kinases, the most studied is the type I PI3K, which can be directly activated by cell surface receptors. Type I PI3K is divided into IA type and IB type according to its activation mode. Class IA PI3K is activated by RTK, G protein-coupled receptors, and small G protein RAS. On the other hand, the IB-type PI3K composed of a single subunit (PI3Kγ) is only activated by G protein-coupled receptors. Class I PI3K is composed of regulators (p85α, p85β, and p85γ) and catalysts (p110α, p110β, p110δ, and p110γ); Among them, p110α and p110β are widely expressed in many tissues, while p110δ and p110γ are mainly expressed in white. blood cells . Activation of ligands (including growth factors) leads to tyrosine phosphorylation of cell surface receptors. The p85 regulatory subunit of IA PI3Ks binds directly to tyrosine receptors on the cell membrane, which are activated by ligands such as platelet growth factor and receptor-related proteins (such as IRS1). Therefore, this binding activates the catalytic subunit p110, which catalyzes the conversion of PIP2 to PIP3. As a second messenger, PIP3 regulates many downstream signaling pathways. Class IB PI3K is a heterodimer composed of the regulatory subunit p101 and the catalytic subunit p110γ. IB-like PI3K is activated by direct binding of p110γ to the Gβγ subunit of the G-protein coupled receptor
As the main molecule downstream of PI3K signaling pathway, serine/threonine protein kinase AKT includes three subtypes of AKT1, AKT2 and AKT3, which are encoded by PKBα, PKBβ and PKBγ, respectively. AKT1 is widely expressed in many tissues, AKT2 is mainly expressed in insulin-sensitive tissues, but at low levels in other tissues, and AKT3 is only expressed in the brain and testis. The specific tissue expression patterns of different subtypes of AKT indicate that they play a key role in maintaining the physiological functions of different tissues or organs..
The PI3KAkt pathway has many after-effects, which must be carefully regulated. Downregulation of the PI3KAKT pathway can achieve the goals: levels of PIP3 and inactivation of the AKT protein. The phosphatase tensin homolog (PTEN) is an important down-regulated protein that can convert PIP3 to PIP2. Protein phosphatase 2A (PP2A) dephosphorylates Akt at Thr308, and PHLPP phosphatase dephosphorylates Akt at Ser473, which are also two negative regulatory proteins. In addition to these regulatory proteins, the pathway itself also has a feedback mechanism: Akt-activated transcription factor NFκB regulates peroxisome proliferator activated receptor agonist delta (PPARβ / δ) and tumor necrosis factor α (TNFα). ), the latter inversely suppresses the expression of PTEN as positive feedback; the negative feedback loop is triggered by mTORC1 and S6K1. S6K1 can phosphorylate IRS1 at multiple serine residues to prevent binding to RTK, thereby inhibiting PI3K activation
Once activated, Akt can move from the plasma membrane to the cytoplasm and then into the nucleus, where many of its substrates reside. Akt phosphorylation can be inhibitory or stimulatory, by inhibiting or enhancing the activity of the target protein. Depending on the target protein, Akt can regulate different cellular functions.