This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
The transforming growth factor beta (TGF-) proteins are cytokines that are required for the development and homeostasis of somatic tissue and regulate tumor progression via their downstream signaling.
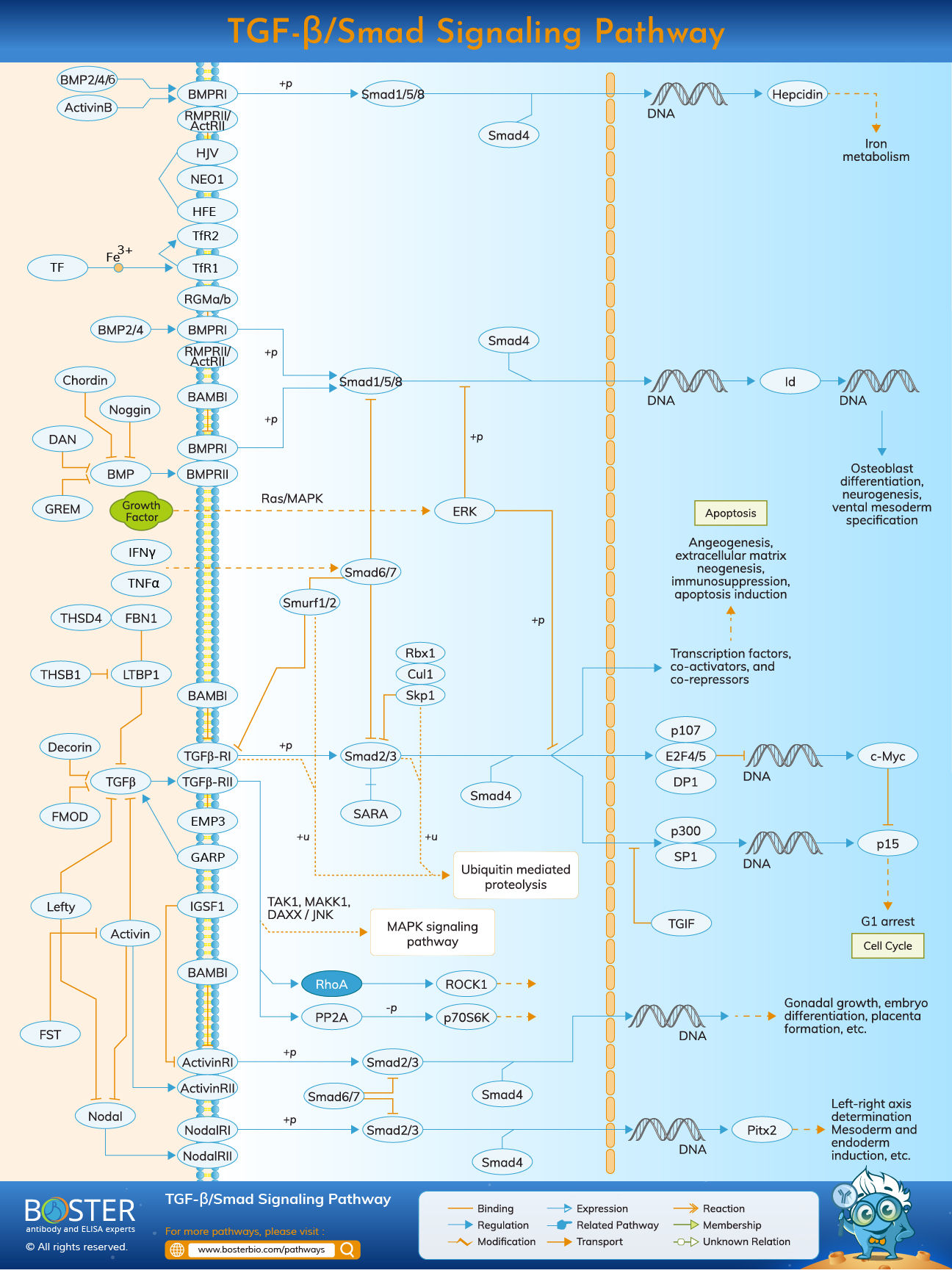
The TGF-β superfamily is comprised of a diverse array of structurally and functionally related proteins, including bone morphogenetic proteins (BMPs), activins, inhibins, growth differentiation factors (GDFs), and glial-derived neurotrophic factors (GDNFs). These members perform multiple functions in a variety of biological processes, including morphogenesis, embryonic development, adult stem cell differentiation, immune regulation, wound healing, inflammation, and cancer. These proteins transmit information by stimulating the formation of heteromeric complexes of serine/threonine kinase receptors of type I and type II. When bioavailable TGF-β binds to a homodimer of TGF-β type II receptors (TRII) on the surface of the target cell, it acts as a growth factor. The TGF-β TRII complex forms a stable complex with a homodimer of the TGF-β type I receptor (TRI). As an active receptor complex, the TRII undergoes autophosphorylation and catalyzes the transphosphorylation of the TRI. The TRI is transphosphorylated to initiate kinase activity.
SMADs were initially identified as the products of the Drosophila Mad and C. elegans Sma genes, which are downstream of these organisms' BMP-analogous ligand-receptor systems. SMADs are expressed ubiquitously throughout development and in all adult tissues, and a large number of them are synthesized from alternatively spliced messenger RNAs. Intracellular TGF-β signaling is mediated by SMAD proteins.
Members of the SMAD family can be classified into three groups based on their functions: 1) the receptor-regulated SMADs (R-SMADs), which include SMAD1, SMAD2, SMAD3, SMAD5, and SMAD8; 2) the common SMADs (Co-SMADs), which include only SMAD4; and 3) the inhibitory SMADs (I-SMADs), which include SMAD6 and SMAD7. R-SMADs interact with membrane-bound serine/threonine receptors and activate them through their kinase activity. Co-SMAD acts as a cofactor, forming a complex with activated R-SMADs that translocates into the nucleus. I-SMADs act in the opposite direction of R-SMADs, inhibiting TGF-superfamily signaling via a variety of mechanisms.
When a ligand binds to the constitutively active type II receptor kinase, it phosphorylates the type I receptor, activating downstream signaling cascades, including the SMAD pathways. By binding to the TRII, TGF-β s, activin, and Nodal initiate intracellular signaling. Then, TGF-β activates the TRI kinase, resulting in the phosphorylation of SMAD2 and SMAD3, while BMPs can signal through SMAD1, 5, and 8. SMAD2 and SMAD3 are then activated and form oligomeric complexes with SMAD4. The signaling of TGF-s, activin, and Nodal via type I receptors is referred to as activin receptor-like kinase (ALK)-4, -5, and -7, respectively. ALK-1, which is preferentially expressed in ECs, however, binds TGF- and activates the SMAD1/5 pathways.
Recently, it was discovered that BMP-9 and BMP-10 bind to ALK-1. Once activated, R-SMADs form a complex with the common mediator SMAD4 (Co-SMAD) and translocate to the nucleus, where SMAD complexes regulate target gene transcription via their interaction with a variety of transcription factors, including the induction of SMAD7.
TGF-β has been shown to activate a variety of non-SMAD downstream signaling pathways, including extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 MAP kinase. TRII and TRI activate the JNK, p38, and inhibitor of kappa B (IkB) kinase (IKK) pathways via TRAF6 and TAK1, respectively, and the ERK pathway via ShcA. TRAF6 also facilitates TACE-mediated cleavage of TRI; the cleaved intracellular domain (ICD) of TRI then associates with transcription factors to activate gene expression. Through Smurf1 recruitment, the TRI can directly phosphorylate PAR6 and degrade RhoA, resulting in the dissociation of tight junctions. Additionally, TGF- activates the RhoA gene during EMT. RhoA, Rac1, and Cdc42 are members of the Rho family of small GTPases that regulate the formation of stress fibers, lamellipodia, and filopodia, respectively. BMP type II receptors bind and activate LIMK1 to inhibit the actin disassembly factor cofilin in non-SMAD signaling.
Additionally, TGF- activates the RhoA gene during EMT. RhoA, Rac1, and Cdc42 are members of the Rho family of small GTPases that regulate the formation of stress fibers, lamellipodia, and filopodia, respectively. BMP type II receptors bind and activate LIMK1 to inhibit the actin disassembly factor cofilin in non-SMAD signaling.