This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
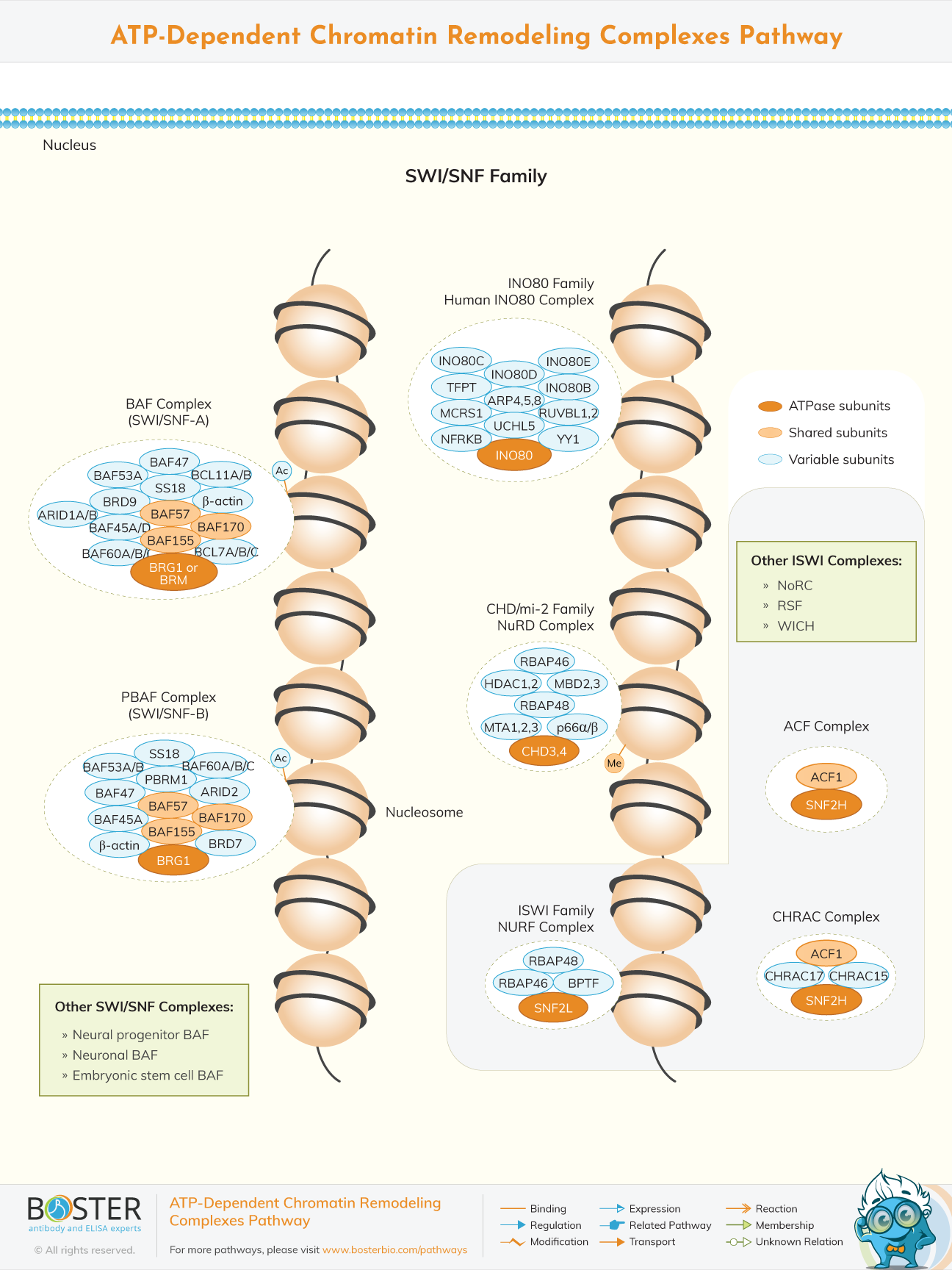
Chromatin remodeling is the dynamic modification of chromatin architecture to allow access of condensed genomic DNA to the regulatory transcription machinery proteins and thereby control gene expression. ATP- dependent remodeling complexes regulate gene expression by either moving, ejecting or restructuring nucleosomes. ATPase domain and energy from hydrolysis of ATP allows these remodeling complexes to reposition nucleosomes along DNA, eject or assemble histones on/off of DNA or facilitate exchange of histone variants, resulting to creation of nucleosome free regions of DNA for gene activation. Many and different people were involved in different discoveries regarding chromatin and its remodeling . Some include (Walter Fleming) who discovered chromatin in 1880,(John,E.W) who discovered The histones, Their interactions with DNA and some aspects of gene control in 1969,(wolffe, A) who discovered chromatin structures and its function.
Many and different people were involved in different discoveries regarding chromatin and its remodeling . Some include (Walter Fleming) who discovered chromatin in 1880,(John,E.W) who discovered The histones, Their interactions with DNA and some aspects of gene control in 1969,(wolffe, A) who discovered chromatin structures and its functions.
Inventory of chromatin remodeling factors started with that of SWI/SNF family. It was originally identified as a regulator of mating type switching (SWI) or as a requirement for growth on energy sources other than sucrose (SNF - sucrose nonfermenting). Human SWI /SNF can be characterized as being of two forms namely BAF (hBRM/BRG1) and PBAF(polybromo associated BAF). It is composed of at least 15 different subunits that invariably include a core ATPase of either BRM or Brg 1 that can provide the necessary energy to the complex of nucleosome remodeling activity. Several common members include BAF 155, BAF 45A/B/C/D,BAF 47, BAF 53A/B,BAF 57, BAF 6A/B/C are shared by the complexes. BRM/BRG1 molecules contain bromodomains that allow binding to acetylated - lysine residues. BAF complexes exists in a wide variety of cell specific, disease specific heterogeneous configuration each containing a total of 12-14 subunits that always include core subunits which are BRM, BRG1, BAF 170, BAF 155, and BAF 47. Complexes with various subunits combinations have been detected in different cell or tissue types during development. Simple example is SWI complex with BAF60C of BAF60 subunit functions in gene transcription in muscles and heart while BAF60A isoform of the complex has a limited role in the tissues. During differentiation of neurons BAF53A is compromised and replaced by BAF53B, indicating the importance of SWI /SNF activity for proper neuron development. BAF45A overexpression improves neural progenitor cell mitosis.
Several long term ostogenic signals directly upregulate the PBAF subunits BAF 180,BAF 200 and BRD 7 in mesenchymal stem cells. Loss of the BAF 200 /BAF 180 /BRD 7, compromised the osteogenenesis and osteolineage gene expression. BRG1, BAF 155 and BAF 60A expressions are largely correlated with programing efficiency of pluripotent stem cells in human population. The SWI family remodelers also participate in DNA double - strand break repair and nucleotide excision repair.
One of two conserved ATPase( SNF2H) or (SNF2L). ISWI complexes have been shown to have specific roles in cell proliferation, differentiation,or mutation. SNF2H containing ISWI complexes mainly participate in early development and progenitor cell proliferation while complexes containing SNF2L are mostly involved in cell differentiation and maturation. ISWI containing chromatin remodeling complex ACF1 is required for nucleosome assembly. SNF2L is also critical for granulosa cell proliferation and differentiation during follicugenesis. BPTF (bromodomain PHD finger transcription factors), one of the NURF subunits is critical for CD 4 or CD 8 single positive cells to differentiate into mature T cells. ISWI family also are mainly involved in organizing nucleosomes into a proper bundle form and create equal spacing between nucleosomes.
BPTF (bromodomain PHD finger transcription factors), one of the NURF subunits is critical for CD 4 or CD 8 single positive cells to differentiate into mature T cells. ISWI family also are mainly involved in organizing nucleosomes into a proper bundle form and create equal spacing between nucleosomes.
t either function alone or cooperate with other proteins to form complexes or constitute a CHD subfamily. Different CHD have distinct roles in early development and cell lineage differentiation. CHD complexes function to regulate transcription or suppression of different genes and induce various lineage differentiation. CHD 3 and 4 are most extensively characterized owing to their role in nucleosome remodeling and complex.
Large, multi-subunit NURD complex contains HDAC1 and 2 protein and combines ATP - dependent chromatin remodeling with histone deacetylase activities to balance both repression and transcriptional activities during embryonic development and cancer.
It exhibit a conserved insertion in ATPase. This is required for interaction between RVB1/RVB2 helicase and these complexes. INO80 subfamily include the 1080 complex while SWR is comprised of P400 and SRCAP. They also participate in DNA double strand break repair and nucleotide excesion repair. INO80 complexes were discovered to include the GLI - Kruppel family zinger transcription factor YYI. Human hINO80 consists of a set of evolutionary conserved subunits which includes :hINO80 snf2 family ATPases, RUVB1/2, actin related proteins (arp 4, arp 5 and arp 8).It also consists of several apparent metazon specific subunits which are :INO80D, INO80E, MCRS1, and NFRKB (nuclear factors related to kB) proteins.
INO80 showcase ATP dependent helicase activity due to presence of Rvb1 and rvb2. INO80 is also critical for meiotic recombination during spermatogenesis.