This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Histone H3 is one of the five major histone proteins that contribute to the structure of chromatin in eukaryotic cells and also plays a critical role in regulating chromatin structure and gene expression
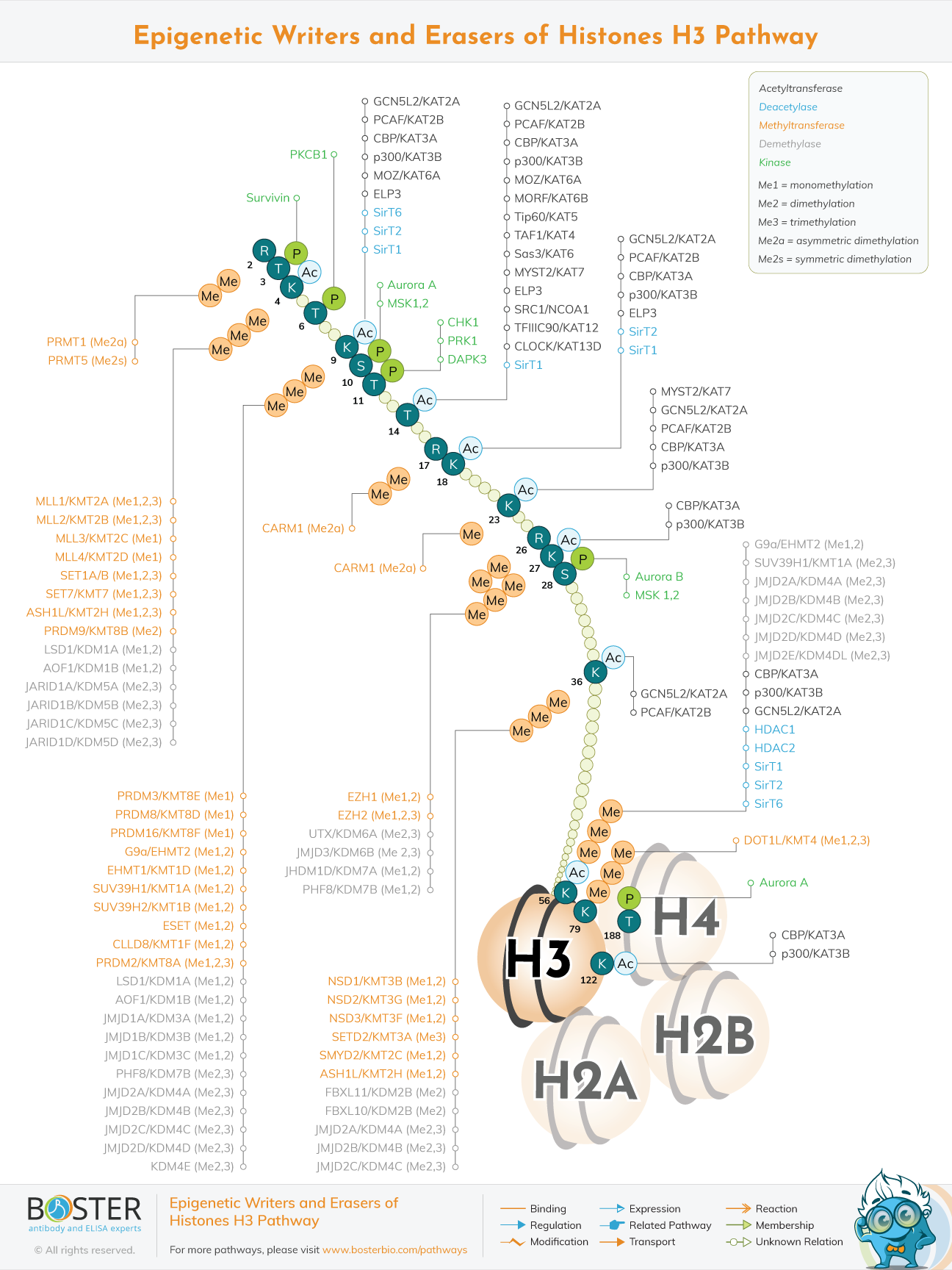
Epigenetic regulatory proteins play a critical role in gene expression regulation by determining whether the DNA sequence is available to the cellular machinery for replication, transcription, DNA repair, recombination, and chromosome segregation. Epigenetic regulation entails multiple classes of enzymes post-translationally modifying amino acid residues within histone proteins (termed writer and eraser proteins). The PTMs in Histone H3 that are involved in the regulation of chromatin structure and gene expression are highlighted in this section. Histone H3 is subjected to a variety of post-translational modifications (PTMs), including acetylation, methylation, and phosphorylation. These PTMs can occur at a variety of histone H3 residues and regulate a variety of processes, including nuclear organization, chromatin structure, and chromatin binding protein recruitment (termed reader proteins).
In comparison to the other four types of histones, histone H3 undergoes only acetylation, methylation, and phosphorylation.These PTMs can occur at a variety of histone H3 residues and regulate a variety of processes involving nuclear organization, chromatin structure, and chromatin binding protein recruitment (also called reader proteins)
Histone lysine acetyltransferases (HATs) catalyze the acetylation of lysine residues, which is reversed by histone deacetylases (HDACs). On histone H3, acetylated lysine residues include Lys4, 9, 14, 18, 23, 27, 36, and 56. Acetylation neutralizes histone H3's positive charge, allowing DNA-binding proteins easier access to the DNA and activating gene expression. Additionally, acetylated lysine residues generate binding domains for bromodomain- and YEATS-containing reader proteins.
Lysine methyltransferases (KMTs) catalyze the methylation of histone lysine and lysine demethylases catalyze the demethylation of histone lysine (KDMs). Lys4, 9, 27, 36, and 79 are methylated lysine residues on histone H3. Each lysine residue can be monomethylated, dimethylated, or trimethylated, and each methylation state appears to have a distinct function. Methylation has no effect on the charge of histones; rather, it regulates the binding of reader proteins and the protein complexes that they associate with. For instance, trimethylation of histone H3 on Lys27 (H3K27me3) by the polycomb repressor complex 2 (PRC2) creates a binding site for the PRC1 complex, both of which contribute to the formation of compact chromatin that is transcriptionally repressive. Alternatively, H3K4me3 is a mark that is recognized by a large number of reader proteins that serve to activate transcription. Methyl-lysine residues serve as binding sites for reader proteins with chromodomains, MBT domains, WD40 domains, and PHD fingers.
Protein arginine methyltransferases (PRMTs) can mono- or di-methylate (symmetrically or asymmetrically) arginine residues, resulting in either gene activation or repression. Additionally, protein arginine deaminase (PADI) proteins can convert methyl-arginine residues to citrulline. Methylation of arginine residues results in the formation of binding sites for reader proteins with Tudor domains, PHD fingers, and WD40 domains.
Finally, kinases phosphorylate histone H3 on serine, threonine, and tyrosine residues, and phosphatases dephosphorylate it. Phosphorylated residues are typically concentrated in the N-terminal tail of histone H3 and, like acetylation, decrease the histone's positive charge. Additionally, phosphorylated residues can serve as binding sites for reader proteins containing 14-3-3 domains or can act as a mask for other reader proteins' binding sites (i.e. HP1 chromodomain protein binding to H3K9Me3 is blocked by H3S10Phos). Along with Thr3, Thr11, and Ser28, phosphorylation of histone H3 Ser10 is primarily associated with chromosome condensation during mitosis and meiosis. Ser10 phosphorylation is frequently used as a marker for cells in mitosis. However, phosphorylation of Ser10 and Ser28 plays a minor role in transcriptional activation, particularly at early genes.