This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
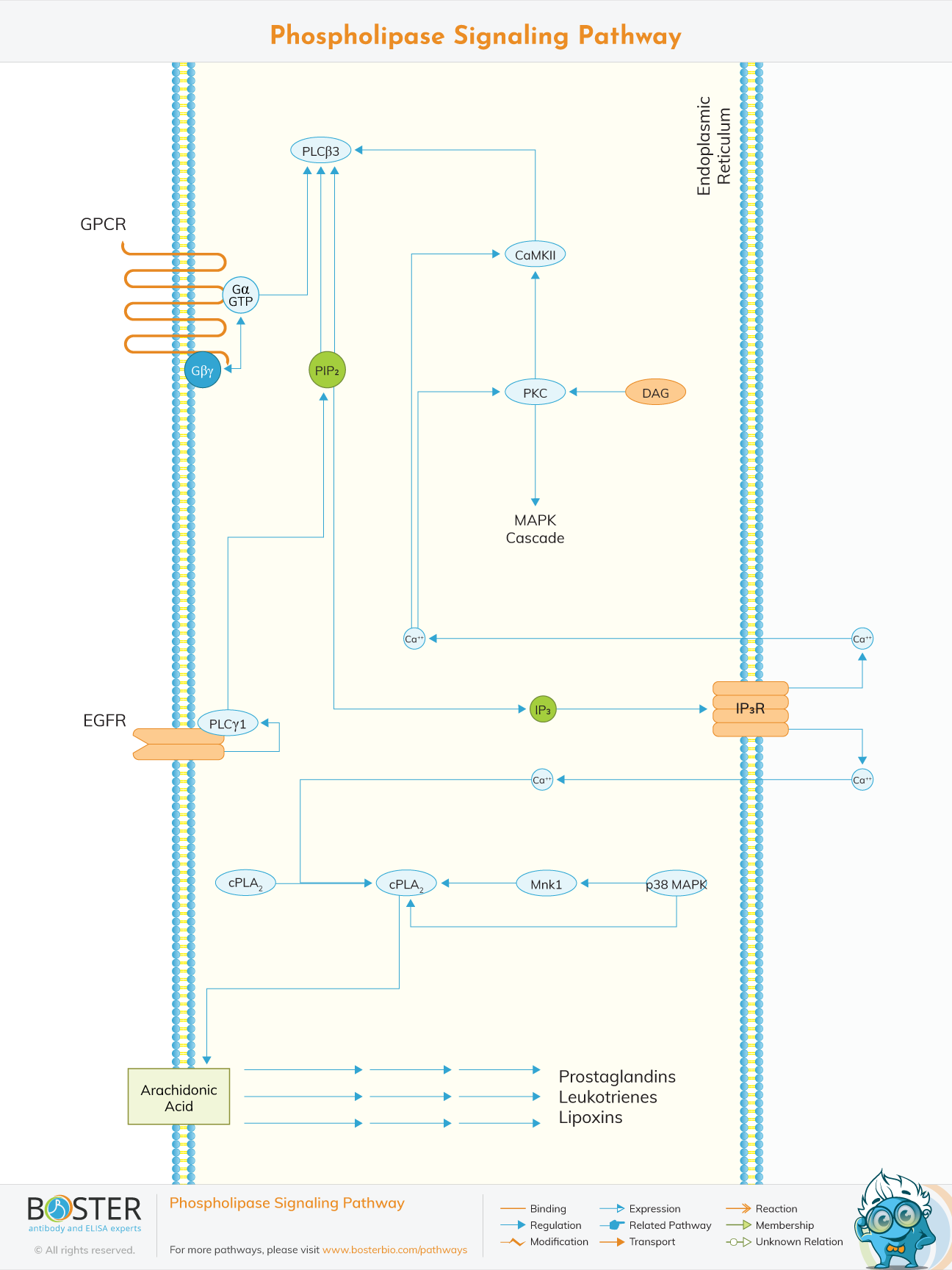
Phospholipase C is an incredibly unique signaling molecule. There is no other enzyme that we are aware of that is capable of directly producing (or modulating) three distinct signals: inositol 1,4,5-trisphosphate (IP3), diacylglycerol, and phosphatidylinositol 4,5-bisphosphate (PIP2). The association of phospholipase C with calcium signaling arguably dates all the way back to Lowell Hokin's (1966) experiments establishing the relative independence of inositol lipid turnover from calcium signaling. This independence from Ca2+ signaling suggested to Bob Michell that inositol lipid turnover, initiated by inositide breakdown via phospholipase C, occurred prior to Ca2+ signaling.
There are numerous mechanisms for Ca2+ signal generation; perhaps the most frequently encountered, and the subject of this review, is one involving a lipid metabolizing enzyme called phospholipase C.
Phosphoinositide-specific phospholipase C (PLC) is involved in transmembrane signaling in a significant way. PLC hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) in response to extracellular stimuli such as hormones, growth factors, and neurotransmitters to produce the secondary messengers inositol 1,4,5-triphosphate (IP3) and diacylglycerol (DAG). PLCs are classified into at least four families: PLC, PLC, PLC, and PLC. Phosphorylation plays a significant role in regulating PLC activity. For example, phosphorylating PLC3 at Ser1105 with PKA or PKC inhibits activity, whereas phosphorylating PLC at Tyr 771, 783, and 1245 with both receptor (EGFR) and nonreceptor tyrosine kinases (Syk) activates it. Additionally, members of the PLC subfamily are activated by heterotrimeric G-protein - or /-subunits and play a critical role in GPCR signaling cascades.
Additionally, members of the PLC subfamily are activated by heterotrimeric G-protein - or /-subunits and play a critical role in GPCR signaling cascades.