This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
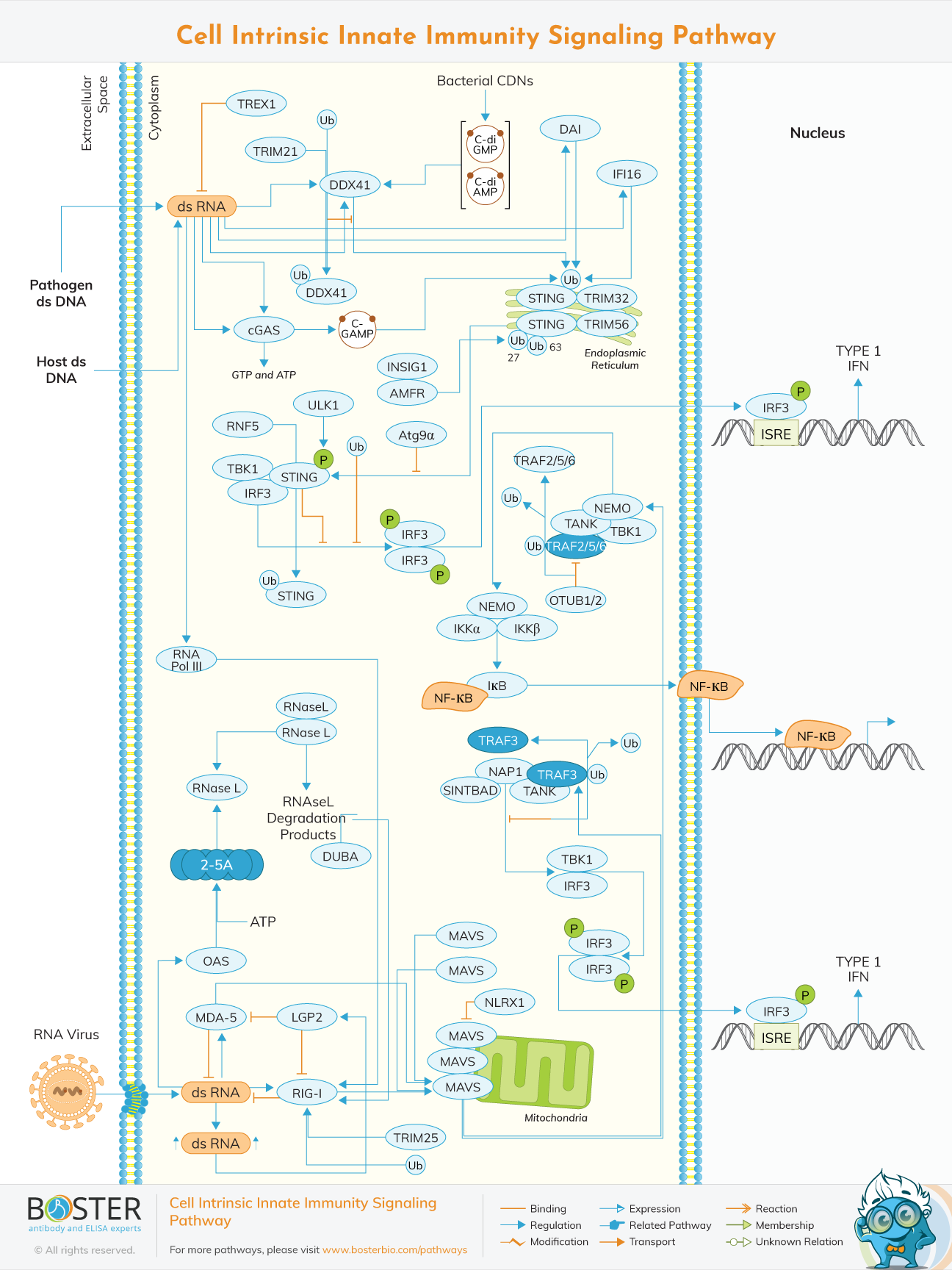
It is a complex cascade that quickly recognizes infections threats through multiple germline-encoded cell surfaces and transmits signals for the deployment of proper countermeasures through the adaptors, kinases, and transcription factors, resulting in the production of cytokines. The innate immune system is an evolutionarily conserved system that senses and defends against infections and irritations. In response to the innate immune system to pathogenic signals, inflammatory responses must be rapid and specific to establishing a physical barrier against the spread of the infection and must be subsequently terminated once the pathogens have been cleared. Intrinsic immunity is conferred by restriction factors that are largely pre-existing in certain cell types. Intrinsic immunity is mediated by constitutively expressed host cell restriction factors that can directly and immediately act to control the viral gene expression.
Many scientists from different corners of the world have dedicated themselves to the research and studies concerning cell intrinsic innate immunity. Some include Tang, E. D, and Wang. C who researched on MAVS self-association mediates antiviral immune signaling in year 2009. Shu, C. Sankaran, B. Charton, C. T Herr in 2003 they based their research and published their studies on structural insight into the functions of TBK1 in innate antimicrobial immunity.
,
The identification of foreign molecular components in the cytosol, such as viral and bacterial nucleic acids, is how infections are detected intracellularly. Through the TBK1-IRF-3 and NF-B pathways, the innate immune system can promote the production of type I interferons (IFNs) and cytokines (TNF-) after it has been recognized. MAVS (mitochondrial adapter protein) and STING (endoplasmic reticulum adapter protein) respond to RNA and DNA sensing pathways, respectively.
RLRs, such as RIG-1 and MDA5, are capable of detecting double-stranded viral RNA and 5'-triphosphate short dsRNA. Through the interaction of common caspase recruitment domains, MDA5 and RIG-1 cause the mitochondrial antiviral-signaling protein (MAVS) to dimerize. Through the interaction of a TRAF-interaction motif and TRAF domains, MAVS dimerization permits TNF receptor-associated factor 3 (TRAF3) binding. The adaptor proteins TANK, NAP1, and SINTBAD are then recruited by TRAF3. TANK connects upstream RLR signaling to the TBK1 (TANK binding kinase 1), which phosphorylates Interferon Regulatory Factor 3. (IRF-3). Phosphorylation and dimerization of IRF-3 causes it to translocate to the nucleus, where it binds to the Interferon-Stimulated Response Element (ISRE), resulting in type I Interferon gene production.
Multiple portals input into the TBK1-IRF-3 signaling pathway from cytosolic double-stranded DNA detection. The cytosolic DNA binding sensor cyclic GMP-AMP synthase (cGAS) produces the cyclic dinucleotide c-GMP-AMP in response to DNA binding (cGAMP). cGAMP binds to STING and stimulates IFN expression through the TBK1-IRF-3 pathway. The STING route is also used by other dsDNA sensors such IFI16, DAI, and DDX41. Furthermore, some pathogenic bacterial infections create cyclic di-nucleotide second messengers that activate the STING pathway. Finally, cytosolic DNA can activate the RIG-I-MAVS pathway by transcribing viral 5′-triphosphate RNA with DNA-dependent RNA polymerase III (RNA Pol III).
NF-B mediated transcription is also induced via the RLR pathway. This occurs in the case of RNA sensing via RLR-induced MAVS polymerization and subsequent binding of TRAF proteins (TRAF2/5/6), whose activity is mediated by NLRX1. TRAF2/5/6 ubiquitination is needed for interaction with MAVS and is inhibited by OTUB1/2 deubiquitination. TRAF2/5/6 activates downstream NF-B signaling by recruiting NEMO and the IB kinases IKK/IKK, which phosphorylate IB.
In the STING and MAVS pathways, ubiquitination plays an important regulatory role. For RNA binding activation, RIG-1 requires K63-ubiquitination by TRIM25, however unanchored chains are also sufficient. TRAF3 is modified with K63-ubiquitin, perhaps autocatalytically, to activate IRF-3 phosphorylation, while TRAF3 degradation induced by DUBA-mediated deubiquitination inhibits IFN production. The extent to which STING is regulated reflects its key role.
The poly-ubiquitination that drives STING translocation from the ER through the Golgi to microsomes, a process negatively controlled by Atg9a, is thought to be mediated by TRIM32 (K63), TRIM56 (K63), and AMFR (K27). RNF5 has also been demonstrated to suppress type I IFN production through the degradation of STING by K48 ubiquitin. Ubiquitination regulates DDX41 degradation through the action of TRIM21. Finally, phosphorylation of STING by the serine-threonine kinase ULK1, which inhibits STING pathway activation, can modulate STING activity.