This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

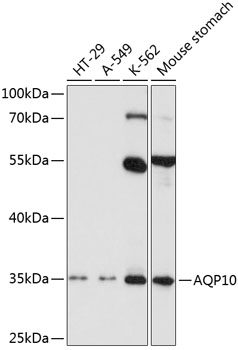
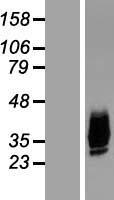
Facts about Aquaporin-10.

Isoform 2 isn't permeable to urea and glycerol. .
| Human | |
|---|---|
| Gene Name: | AQP10 |
| Uniprot: | Q96PS8 |
| Entrez: | 89872 |

| Belongs to: |
|---|
| MIP/aquaporin (TC 1.A.8) family |

AQP-10; AQPA_HUMAN; aquaglyceroporin-10; aquaporin 10; aquaporin-10; Small intestine aquaporin
Mass (kDA):
31.763 kDA

| Human | |
|---|---|
| Location: | 1q21.3 |
| Sequence: | 1; NC_000001.11 (154321059..154325325) |
Expressed exclusively in duodenum and jejunum. Highest expression in absorptive epithelial cells at the tips of villi in the jejunum.
Apical cell membrane; Multi-pass membrane protein. Cell membrane; Multi-pass membrane protein. Lipid droplet. Detected around lipid droplets.




PMID: 11573934 by Hatakeyama S., et al. Cloning of a new aquaporin (AQP10) abundantly expressed in duodenum and jejunum.
PMID: 12084581 by Ishibashi K., et al. Cloning and identification of a new member of water channel (AQP10) as an aquaglyceroporin.