This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

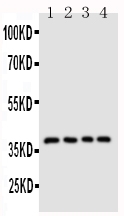
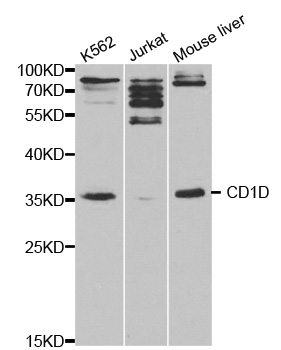
Facts about Antigen-presenting glycoprotein CD1d.

| Human | |
|---|---|
| Gene Name: | CD1D |
| Uniprot: | P15813 |
| Entrez: | 912 |

| Belongs to: |
|---|
| No superfamily |

antigen-presenting glycoprotein CD1d; CD1A; CD1d antigen; CD1D antigen, d polypeptide; CD1d molecule; CD1d; differentiation antigen CD1-alpha-3; HMC class I antigen-like glycoprotein CD1D; MGC34622; R3; R3G1; T-cell surface glycoprotein CD1d; thymocyte antigen CD1D
Mass (kDA):
37.717 kDA

| Human | |
|---|---|
| Location: | 1q23.1 |
| Sequence: | 1; NC_000001.11 (158178030..158186427) |
Expressed on cortical thymocytes, on certain T-cell leukemias, and in various other tissues.
Cell membrane; Single-pass type I membrane protein. Basolateral cell membrane; Single-pass type I membrane protein. Endosome membrane; Single-pass type I membrane protein. Lysosome membrane; Single-pass type I membrane protein. Endoplasmic reticulum membrane; Single-pass type I membrane protein. Subject to intracellular trafficking between the cell membrane, endosomes and lysosomes.




PMID: 2467814 by Calabi F., et al. Two classes of CD1 genes.
PMID: 2463622 by Balk S.P., et al. Isolation and characterization of a cDNA and gene coding for a fourth CD1 molecule.