This website uses cookies to ensure you get the best experience on our website.
- Table of Contents


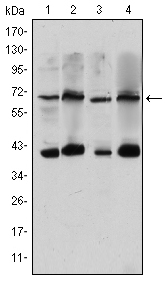
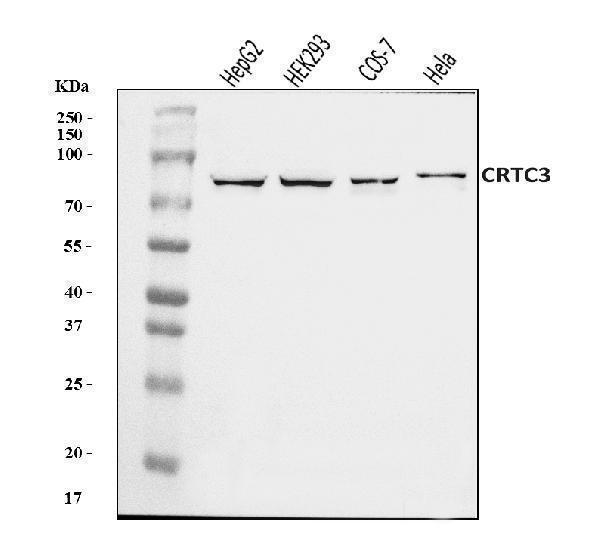
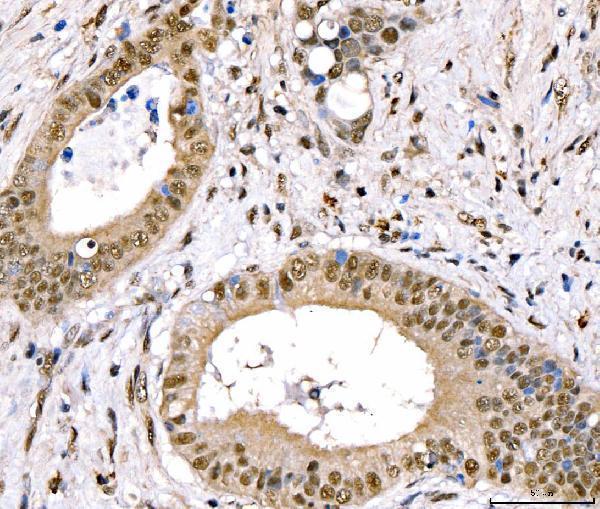
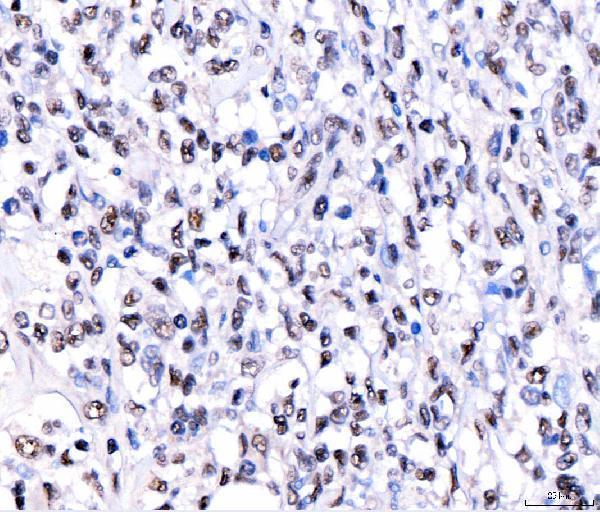
Facts about CREB-regulated transcription coactivator 3.

Enhances the interaction of CREB1 with TAF4. Regulates the expression of specific CREB-activated genes such as the steroidogenic gene, StAR.
| Human | |
|---|---|
| Gene Name: | CRTC3 |
| Uniprot: | Q6UUV7 |
| Entrez: | 64784 |

| Belongs to: |
|---|
| TORC family |

CREB regulated transcription coactivator 3; CRTC3; FLJ21868; TORC3; TORC-3; TORC3CREB-regulated transcription coactivator 3; Transducer of CREB protein 3; transducer of regulated cAMP response element-binding protein (CREB) 3; Transducer of regulated cAMP response element-binding protein 3; transducer of regulated CREB protein 3
Mass (kDA):
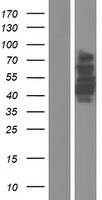
66.959 kDA

| Human | |
|---|---|
| Location: | 15q26.1 |
| Sequence: | 15; NC_000015.10 (90529915..90645345) |
Predominantly expressed in B and T lymphocytes. Highest levels in lung. Also expressed in brain, colon, heart, kidney, ovary, and prostate. Weak expression in liver, pancreas, muscle, small intestine, spleen and stomach.
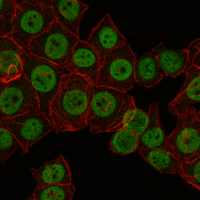
Nucleus. Cytoplasm. Appears to be mainly nuclear (PubMed:15454081). Translocates to the nucleus following adenylyl cyclase or MAP kinase activation (PubMed:30611118).



PMID: 14506290 by Iourgenko V., et al. Identification of a family of cAMP response element-binding protein coactivators by genome-scale functional analysis in mammalian cells.
PMID: 12693554 by Jikuya H., et al. Characterization of long cDNA clones from human adult spleen. II. The complete sequences of 81 cDNA clones.