This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

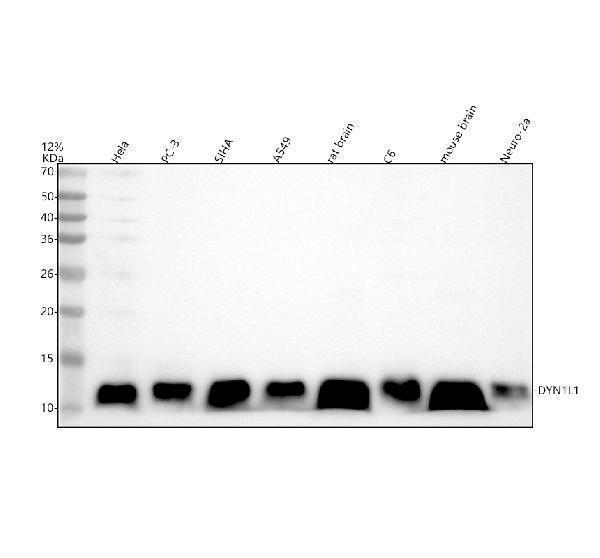
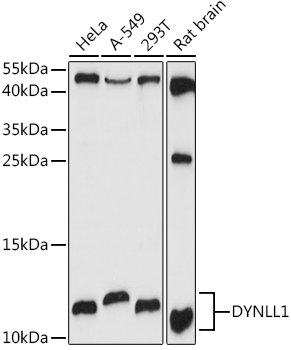
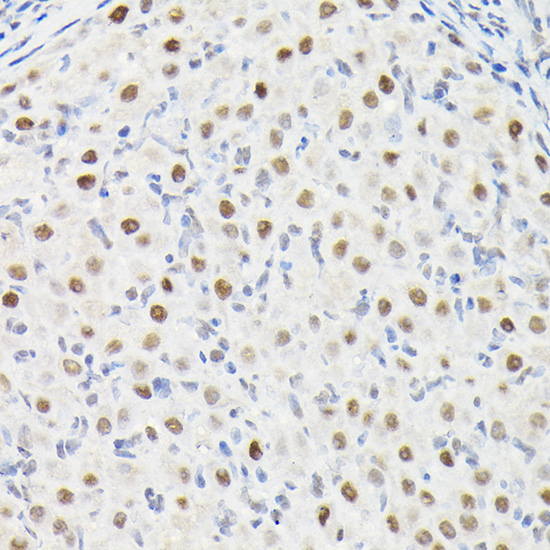
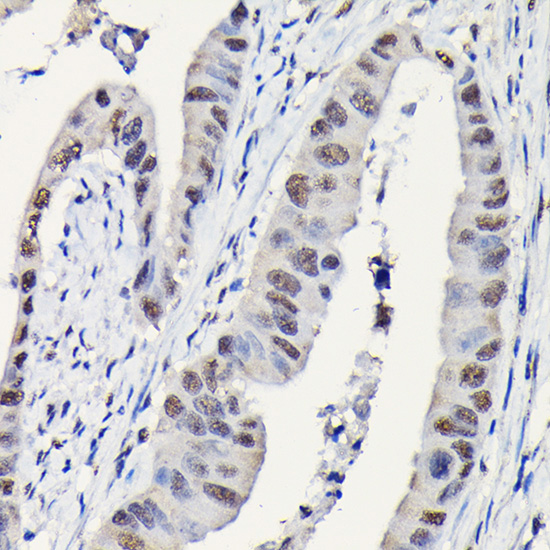
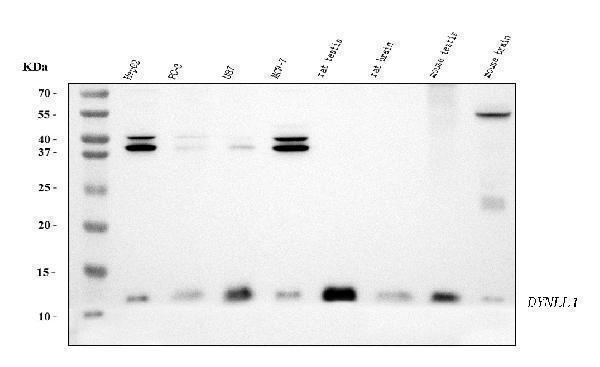
Facts about Dynein light chain 1, cytoplasmic.

May play a role in changing or maintaining the spatial distribution of cytoskeletal structures. .
| Human | |
|---|---|
| Gene Name: | DYNLL1 |
| Uniprot: | P63167 |
| Entrez: | 8655 |

| Belongs to: |
|---|
| dynein light chain family |

cytoplasmic dynein light polypeptide; DLC1; DLC1DNCLC1; DLC8; DLC8MGC126137; DNCL1; DNCL1MGC126138; DNCLC1; dynein light chain 1, cytoplasmic; Dynein light chain LC8-type 1; dynein, cytoplasmic, light polypeptide 1; dynein, light chain, LC8-type 1,8 kDa dynein light chain; DYNLL1; hdlc1; LC8; LC8a; PIN; PINLC8a; Protein inhibitor of neuronal nitric oxide synthase
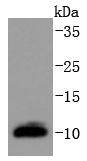
Mass (kDA):
10.366 kDA

| Human | |
|---|---|
| Location: | 12q24.31 |
| Sequence: | 12; NC_000012.12 (120469842..120498493) |
Ubiquitous.
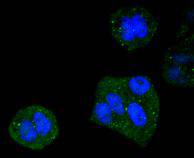
Cytoplasm, cytoskeleton, microtubule organizing center, centrosome. Cytoplasm, cytoskeleton. Nucleus. Mitochondrion. Upon induction of apoptosis translocates together with BCL2L11 to mitochondria.




PMID: 8628263 by Dick T., et al. Cytoplasmic dynein (ddlc1) mutations cause morphogenetic defects and apoptotic cell death in Drosophila melanogaster.
PMID: 10198631 by Puthalakath H., et al. The proapoptotic activity of the Bcl-2 family member Bim is regulated by interaction with the dynein motor complex.