This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

3 Citations 4 Q&As
Facts about Corticosteroid 11-beta-dehydrogenase isozyme 2.

.
| Human | |
|---|---|
| Gene Name: | HSD11B2 |
| Uniprot: | P80365 |
| Entrez: | 3291 |

| Belongs to: |
|---|
| short-chain dehydrogenases/reductases (SDR) family |

11 betaHSD2; 11 beta-HSD2; AME; corticosteroid 11-beta-dehydrogenase isozyme 2,11-beta-HSD2; EC 1.1.1; EC 1.1.1.-; HSD11B2; HSD11KAME1; HSD2,11-DH2; hydroxysteroid (11-beta) dehydrogenase 2; NAD-dependent 11-beta-hydroxysteroid dehydrogenase; SDR9C3; short chain dehydrogenase/reductase family 9C member 3,11-beta-hydroxysteroid dehydrogenase type 2
Mass (kDA):
44.127 kDA

| Human | |
|---|---|
| Location: | 16q22.1 |
| Sequence: | 16; NC_000016.10 (67431121..67437553) |
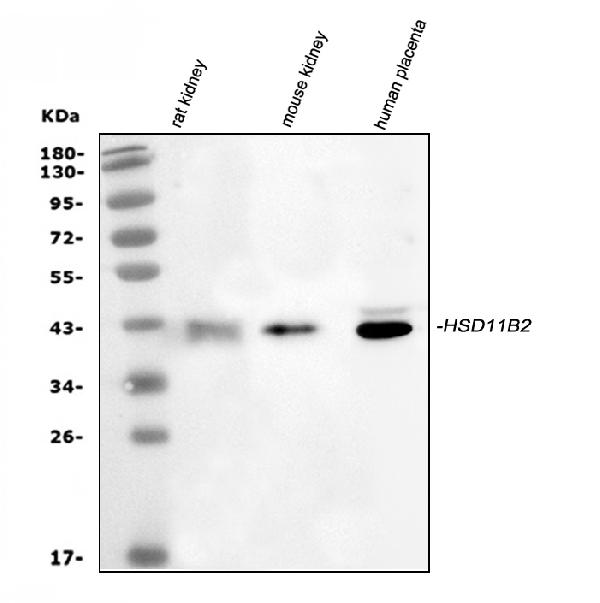
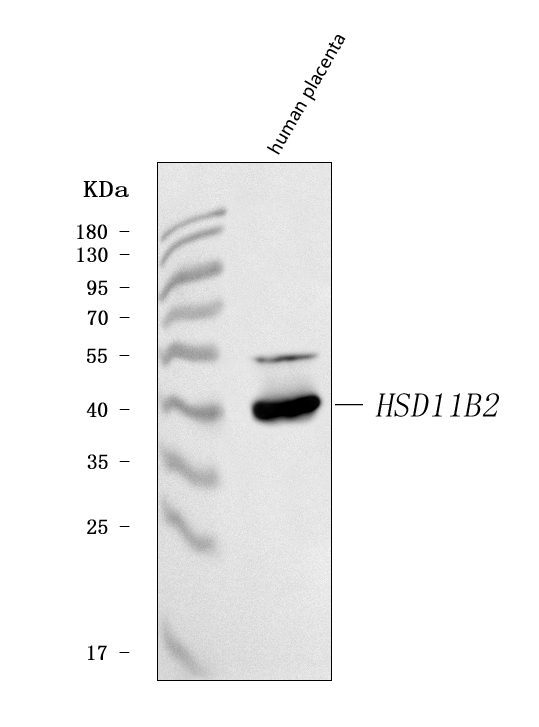
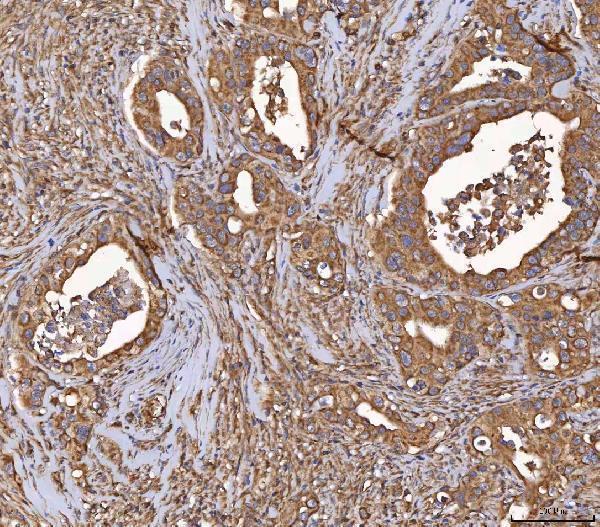
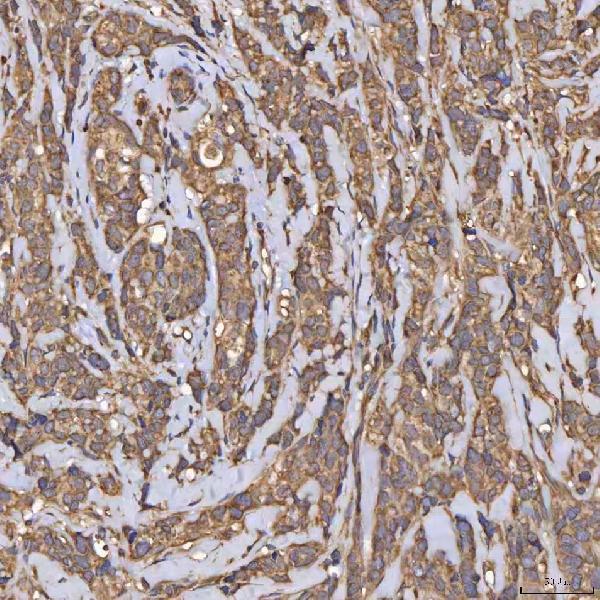
Expressed in kidney, pancreas, prostate, ovary, small intestine and colon. At midgestation, expressed at high levels in placenta and in fetal kidney and, at much lower levels, in fetal lung and testis (PubMed:8530071).
Microsome. Endoplasmic reticulum.




PMID: 7859916 by Albiston A.L., et al. Cloning and tissue distribution of the human 11 beta-hydroxysteroid dehydrogenase type 2 enzyme.
PMID: 8530071 by Agarwal A.K., et al. Gene structure and chromosomal localization of the human HSD11K gene encoding the kidney (type 2) isozyme of 11 beta-hydroxysteroid dehydrogenase.