This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

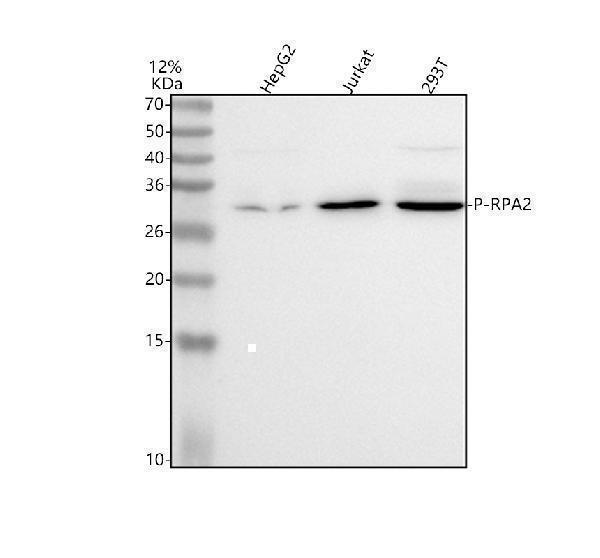




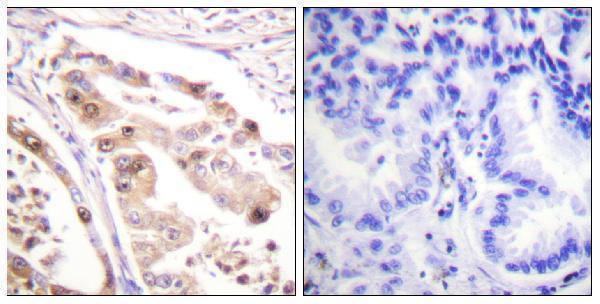
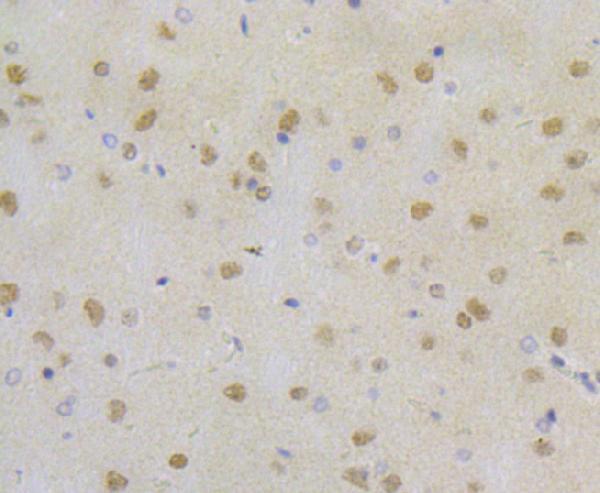
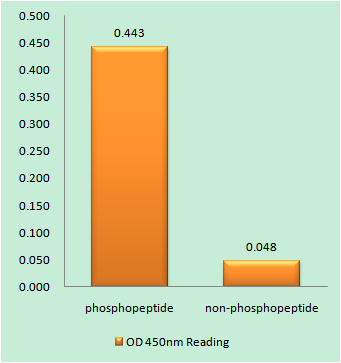
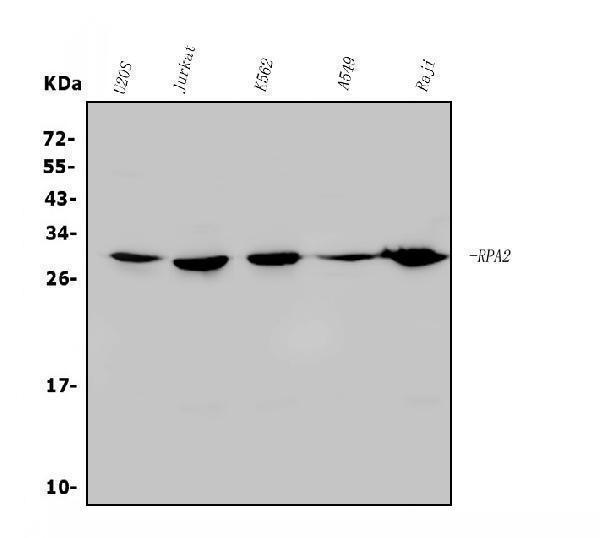
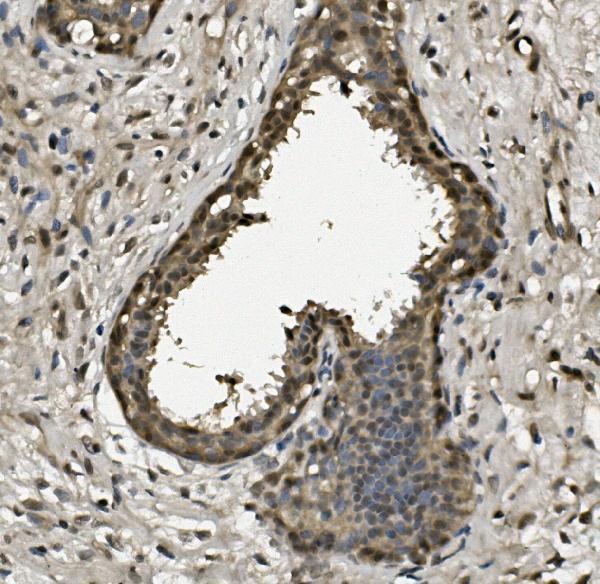
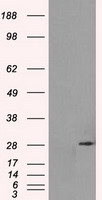
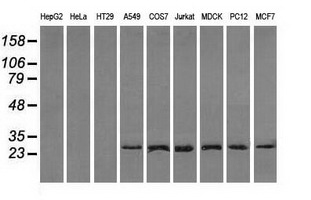
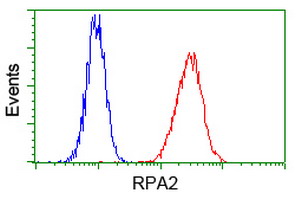
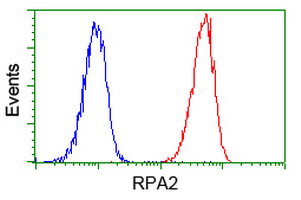
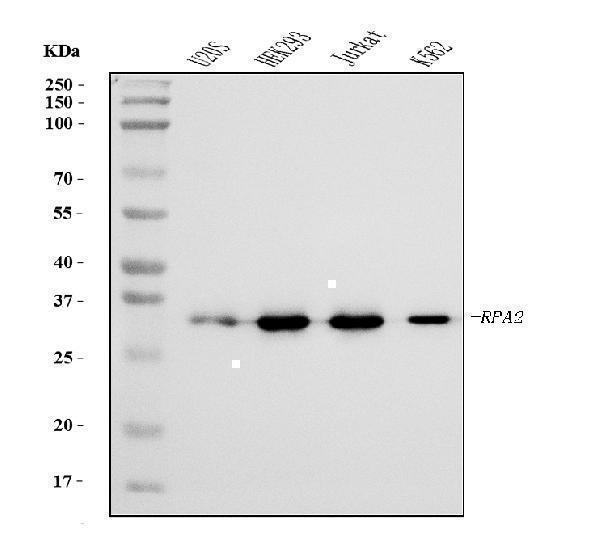
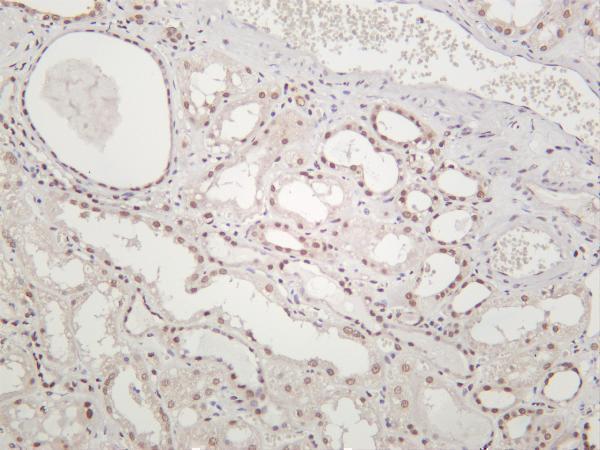
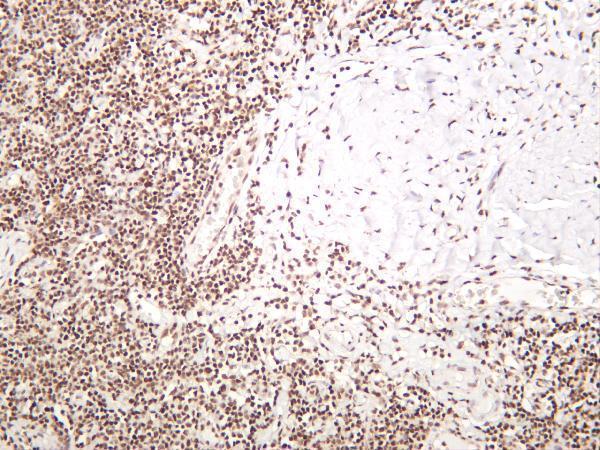
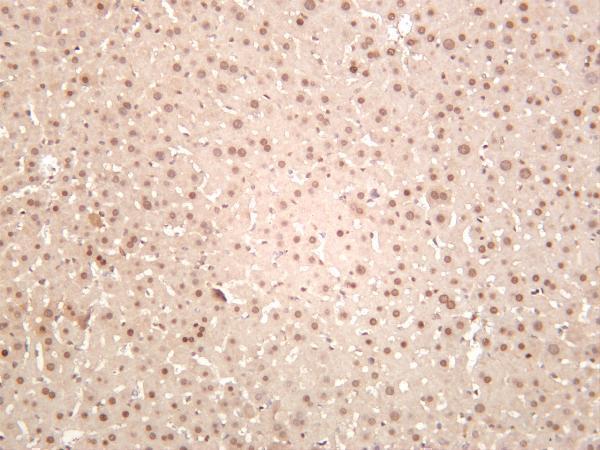
Facts about Replication protein A 32 kDa subunit.

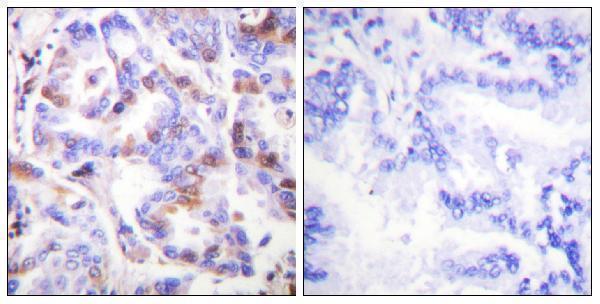
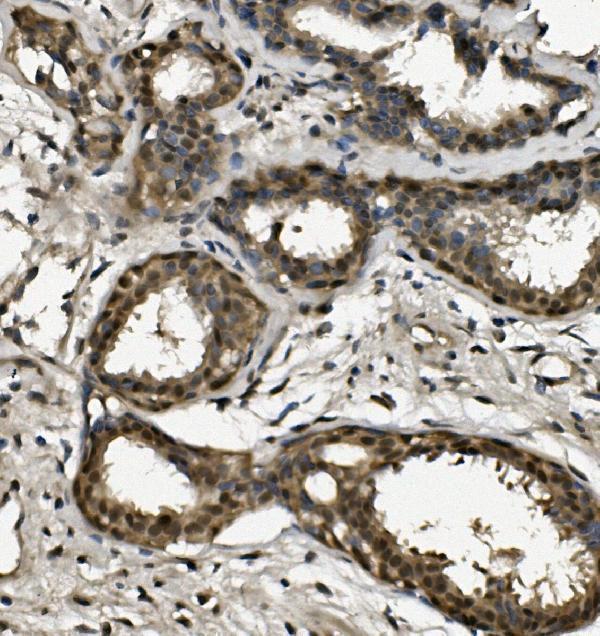
Thereby, it plays an essential role both in DNA replication and the cellular response to DNA damage. From the cellular response to DNA damage, the RPA complex controls DNA repair and DNA damage checkpoint activation.
| Human | |
|---|---|
| Gene Name: | RPA2 |
| Uniprot: | P15927 |
| Entrez: | 6118 |

| Belongs to: |
|---|
| replication factor A protein 2 family |

REPA2; Replication factor A protein 2; replication protein A 32 kDa subunit; Replication protein A 34 kDa subunit; replication protein A2 (32kD); replication protein A2, 32kDa; RF-A protein 2; RP-A p32; RP-A p34; RPA2; RPA32; RPA34
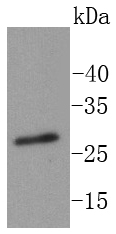
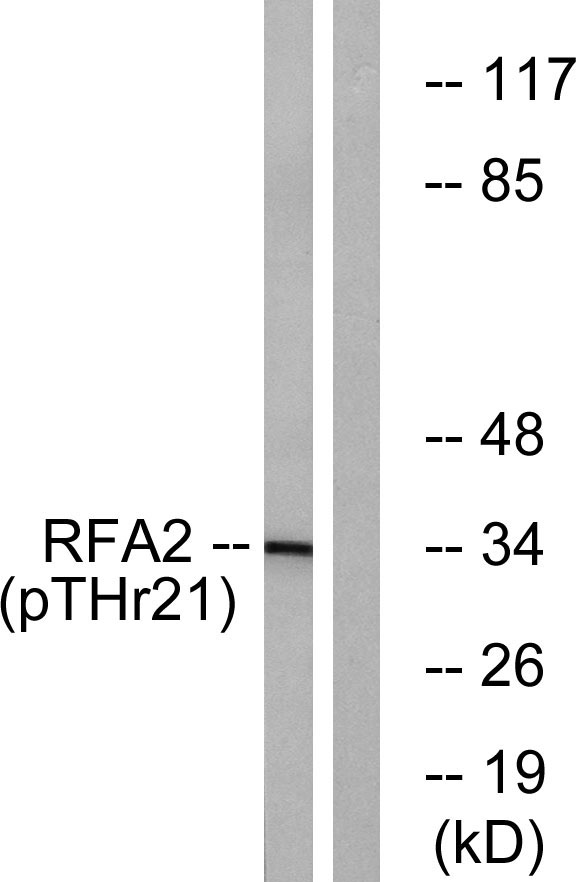
Mass (kDA):
29.247 kDA

| Human | |
|---|---|
| Location: | 1p35.3 |
| Sequence: | 1; NC_000001.11 (27891524..27914797, complement) |
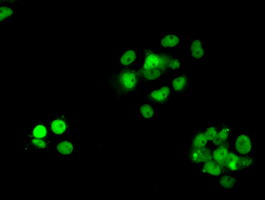
Nucleus. Nucleus, PML body. Redistributes to discrete nuclear foci upon DNA damage in an ATR-dependent manner.





PMID: 2406247 by Erdile L.F., et al. The primary structure of the 32-kDa subunit of human replication protein A.
PMID: 2200738 by Din S., et al. Cell-cycle-regulated phosphorylation of DNA replication factor A from human and yeast cells.