This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

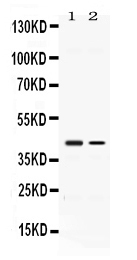
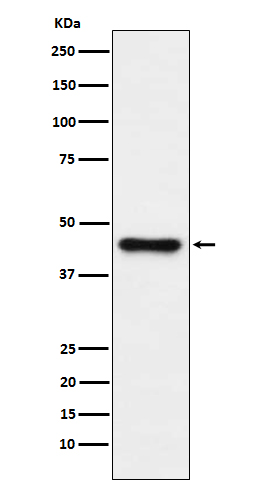
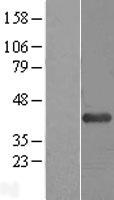
Facts about 1-acylglycerol-3-phosphate O-acyltransferase ABHD5.

Involved in keratinocyte differentiation (PubMed:18832586). Regulates lipid droplet fusion (By similarity).
| Human | |
|---|---|
| Gene Name: | ABHD5 |
| Uniprot: | Q8WTS1 |
| Entrez: | 51099 |

| Belongs to: |
|---|
| peptidase S33 family |

abhydrolase domain containing 5; Abhydrolase domain-containing protein 5,1-acylglycerol-3-phosphate O-acyltransferase ABHD5; CDS; CGI58; EC 2.3.1.51; IECN2; Lipid droplet-binding protein CGI-58; MGC8731; NCIE2CGI-58
Mass (kDA):
39.096 kDA

| Human | |
|---|---|
| Location: | 3p21.33 |
| Sequence: | 3; NC_000003.12 (43690870..43734371) |
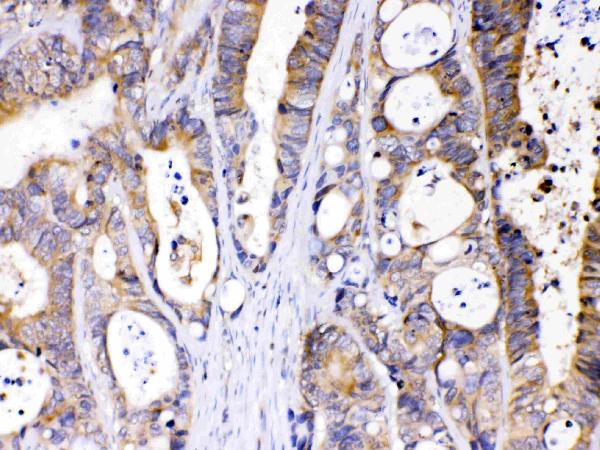
Widely expressed in various tissues, including lymphocytes, liver, skeletal muscle and brain. Expressed by upper epidermal layers and dermal fibroblasts in skin, hepatocytes and neurons (at protein level).
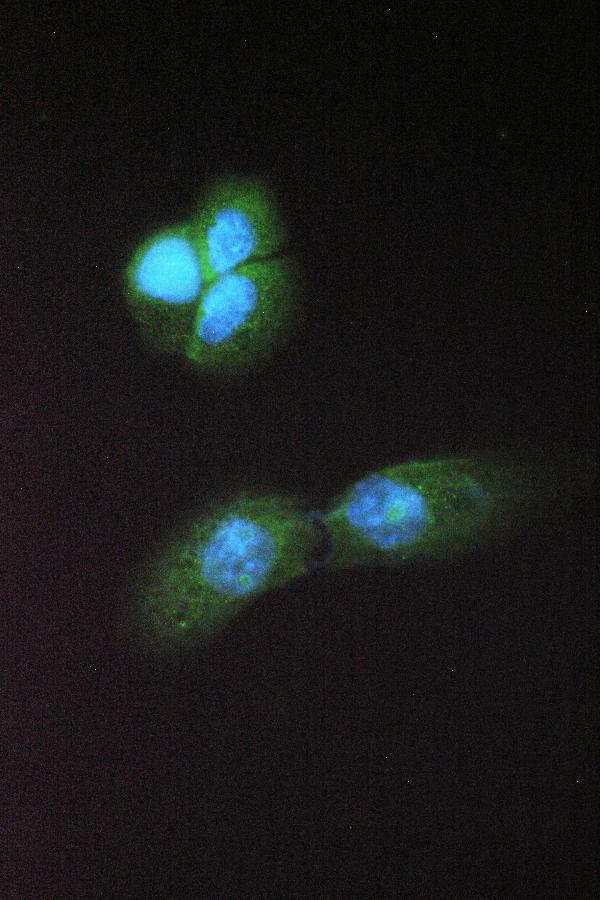
Cytoplasm. Lipid droplet. Cytoplasm, cytosol. Colocalized with PLIN and ADRP on the surface of lipid droplets. The localization is dependent upon the metabolic status of the adipocytes and the activity of PKA (By similarity).




PMID: 11590543 by Lefevre C., et al. Mutations in CGI-58, the gene encoding a new protein of the esterase/lipase/thioesterase subfamily, in Chanarin-Dorfman syndrome.
PMID: 16679289 by Lass A., et al. Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by CGI-58 and defective in Chanarin-Dorfman Syndrome.