This website uses cookies to ensure you get the best experience on our website.
- Table of Contents




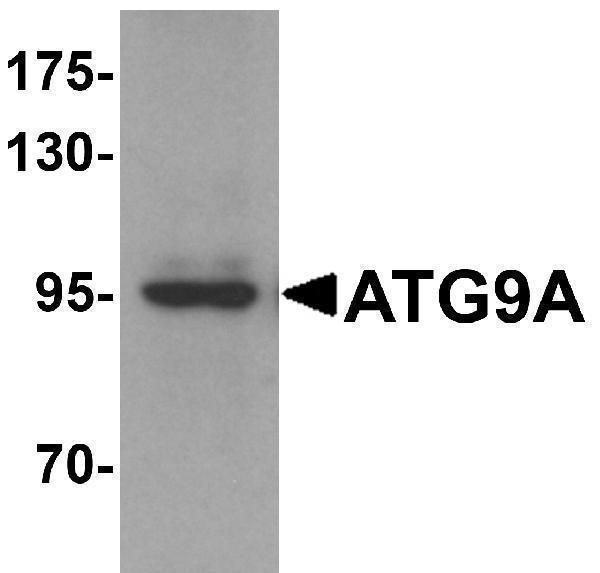
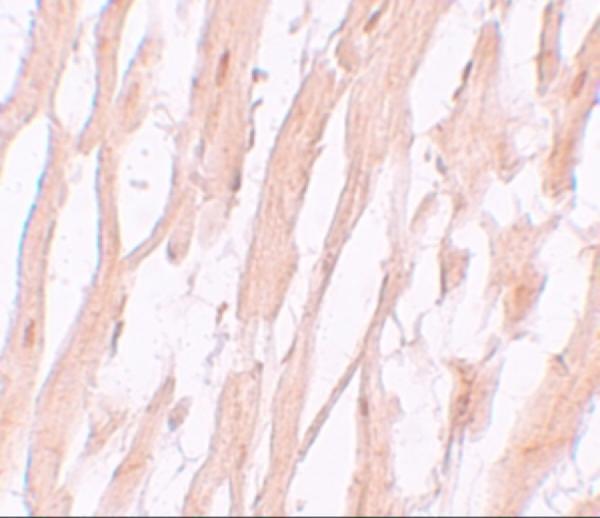
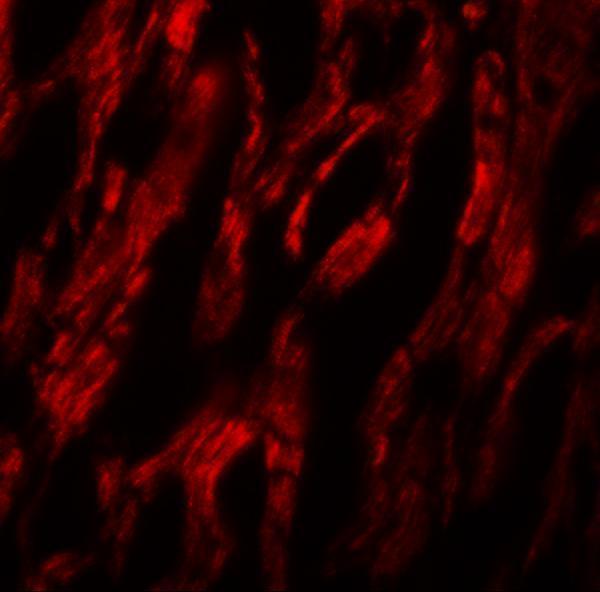
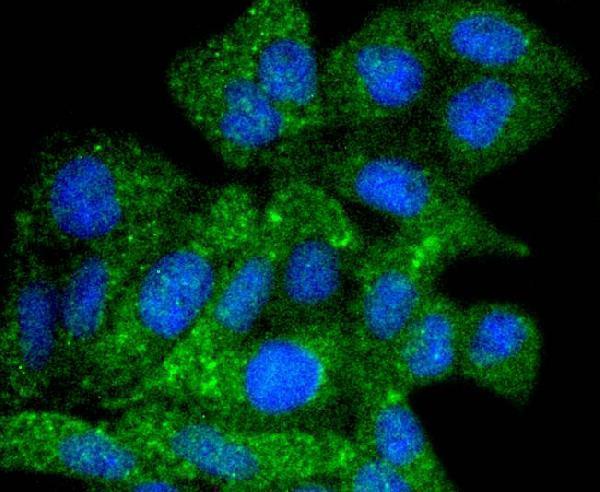
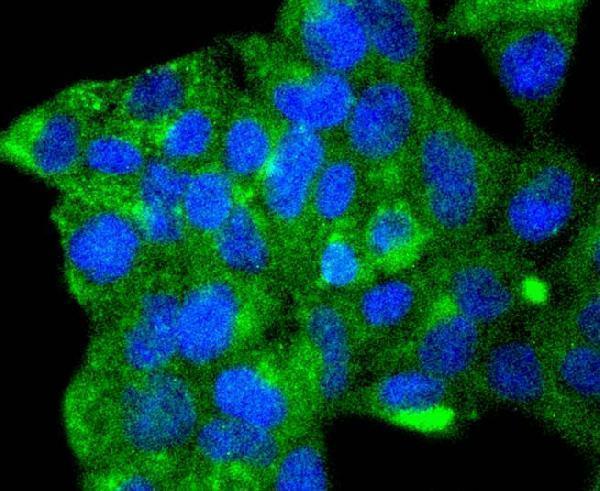
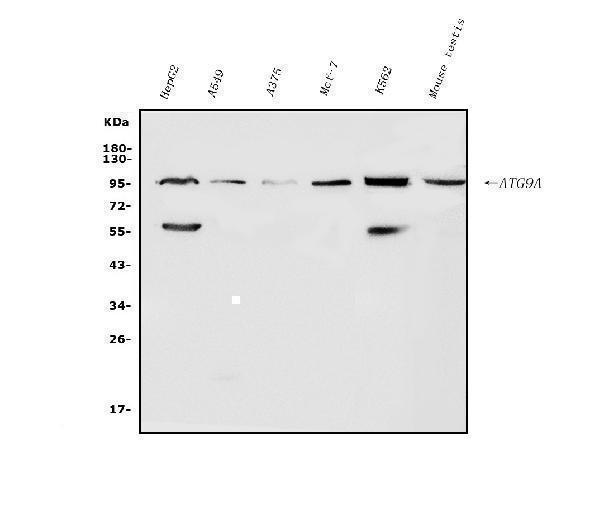
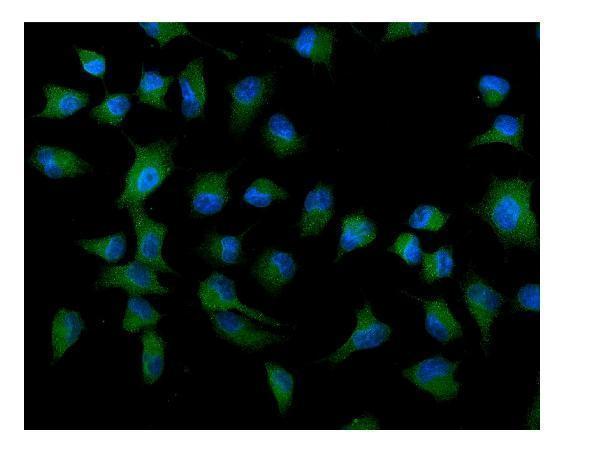
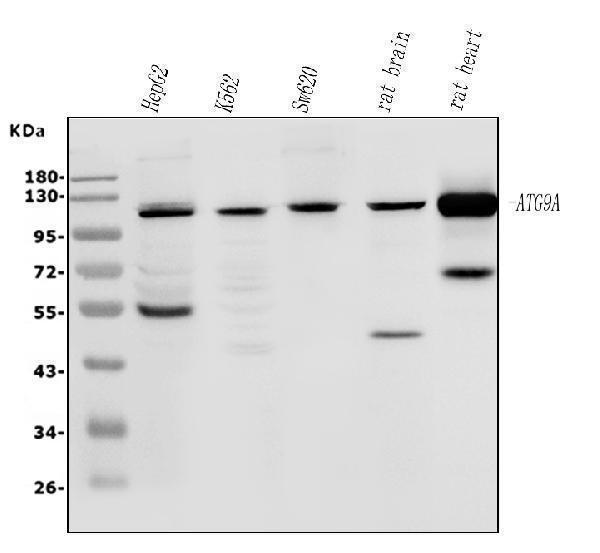
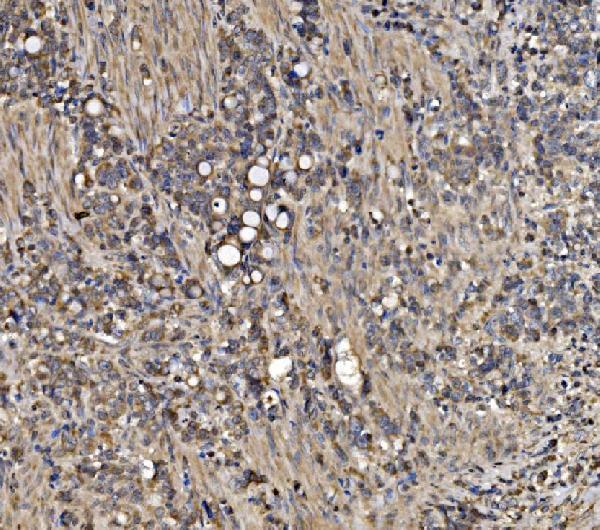
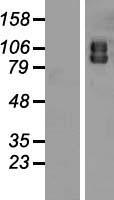
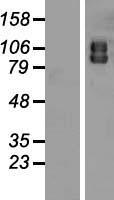
Facts about Autophagy-related protein 9A.

Cycles between a juxta-nuclear trans-Golgi network compartment and late endosomes. Nutrient starvation induces accumulation on autophagosomes.
| Human | |
|---|---|
| Gene Name: | ATG9A |
| Uniprot: | Q7Z3C6 |
| Entrez: | 79065 |

| Belongs to: |
|---|
| ATG9 family |

APG9 autophagy 9-like 1 (S. cerevisiae); APG9 autophagy 9-like 1; APG9L1APG9-like 1; ATG9 autophagy related 9 homolog A (S. cerevisiae); autophagy 9-like 1 protein; autophagy-related protein 9A; FLJ22169; mATG9; MGD3208
Mass (kDA):
94.447 kDA

| Human | |
|---|---|
| Location: | 2q35 |
| Sequence: | 2; NC_000002.12 (219219380..219229636, complement) |
Cytoplasmic vesicle, autophagosome membrane; Multi-pass membrane protein. Golgi apparatus, trans-Golgi network membrane; Multi-pass membrane protein. Late endosome membrane; Multi-pass membrane protein. Endoplasmic reticulum membrane; Multi-pass membrane protein. Under amino acid starvation or rapamycin treatment, redistributes from a juxtanuclear clustered pool to a dispersed peripheral cytosolic pool. The starvation-induced redistribution depends on ULK1, ATG13, as well as SH3GLB1.





PMID: 15755735 by Yamada T., et al. Endothelial nitric-oxide synthase antisense (NOS3AS) gene encodes an autophagy-related protein (APG9-like2) highly expressed in trophoblast.
PMID: 16940348 by Young A.R., et al. Starvation and ULK1-dependent cycling of mammalian Atg9 between the TGN and endosomes.