This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

1 Citations 1 Q&As
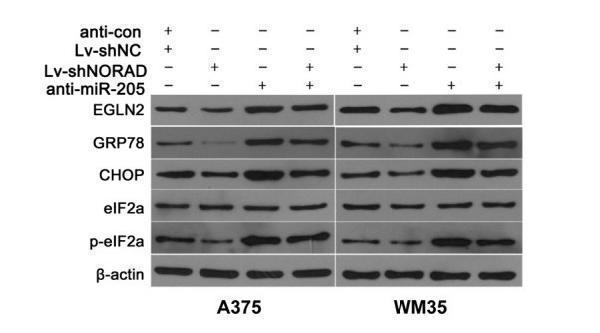
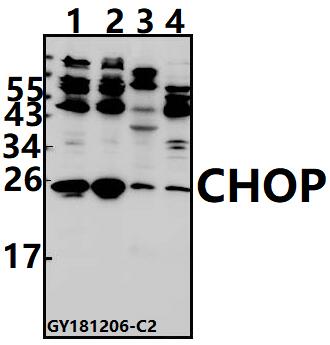
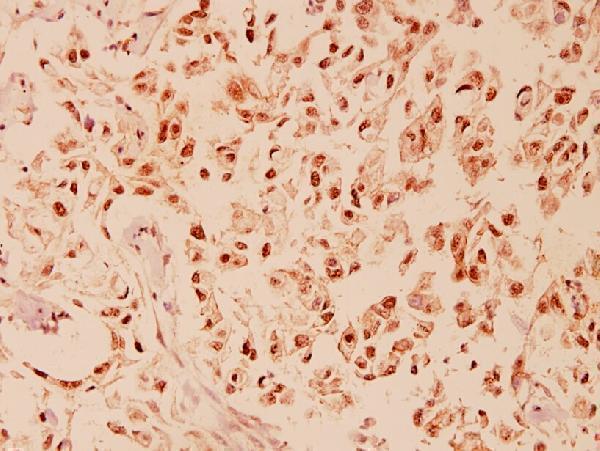
9 Citations 3 Q&As


2 Citations
6 Citations
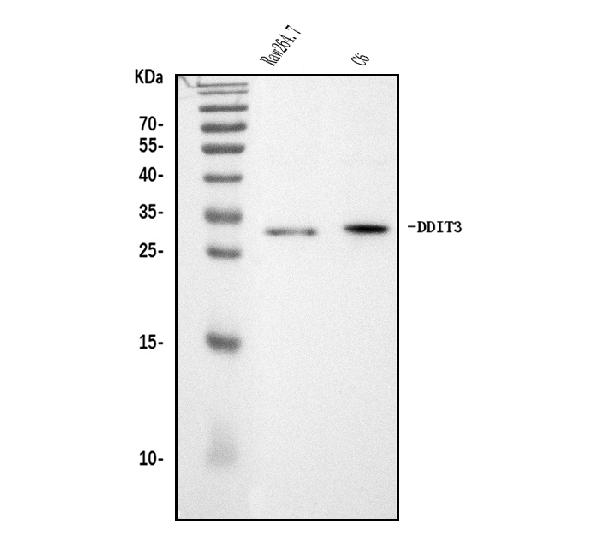
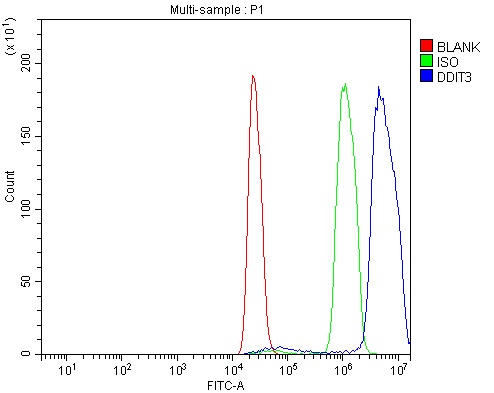
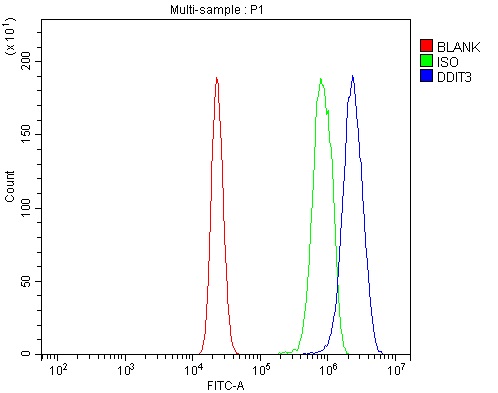
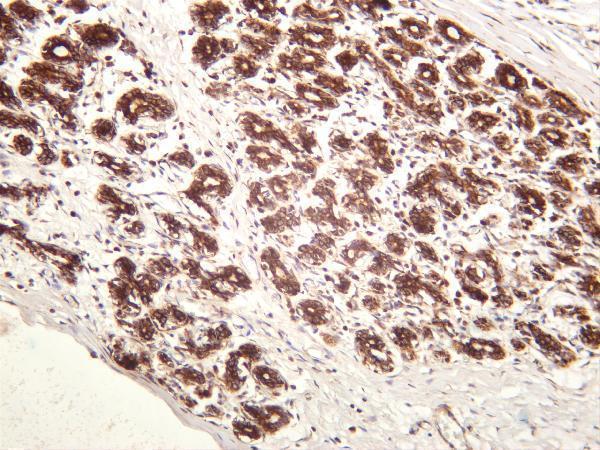
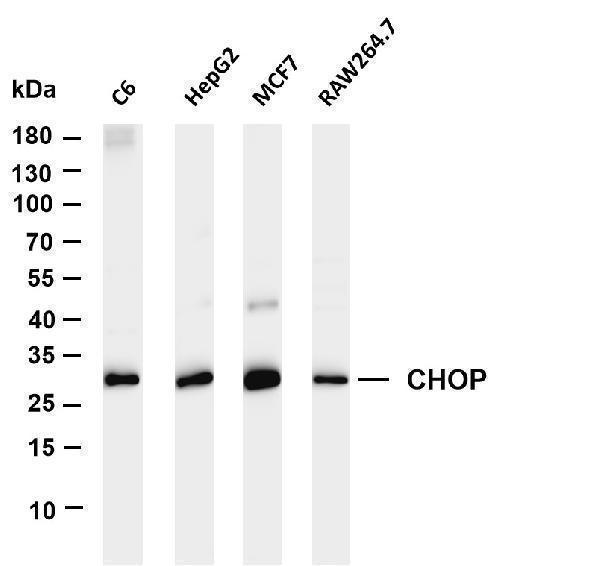
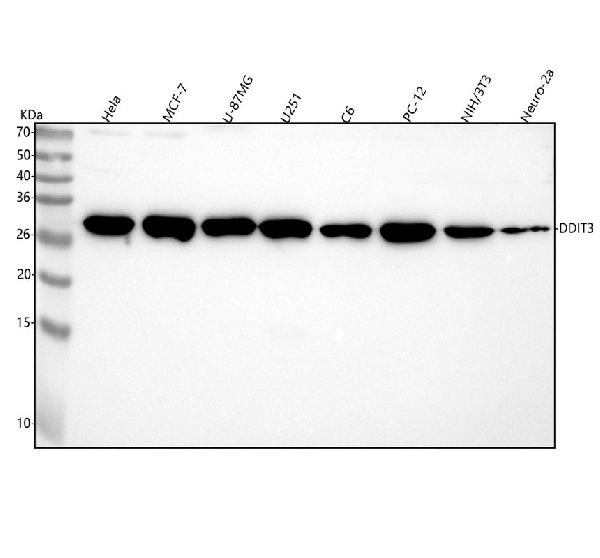
Facts about DDIT3 upstream open reading frame protein.

Acts as a dominant-negative regulator of C/EBP-induced transcription: dimerizes with members of their C/EBP family, impairs their affiliation with C/EBP binding sites in the promoter regions, and inhibits the expression of C/EBP regulated genes. Positively regulates the transcription of TRIB3, IL6, IL8, IL23, TNFRSF10B/DR5, PPP1R15A/GADD34, BBC3/PUMA, BCL2L11/BIM and ERO1L.
| Human | |
|---|---|
| Gene Name: | DDIT3 |
| Uniprot: | P0DPQ6 |
| Entrez: | 1649 |

| Belongs to: |
|---|
| No superfamily |

C/EBP-homologous protein 10; C/EBP-homologous protein; CCAAT/enhancer-binding protein homologous protein; CEBP zeta; CHOP; CHOP10 CEBPZ; CHOP10; CHOP-10; CHOPC/EBP zeta; DDIT3; DDIT-3; DNA damage-inducible transcript 3 protein; DNA-damage-inducible transcript 3; GADD153; Growth arrest and DNA damage-inducible protein GADD153
Mass (kDA):
4.284 kDA

| Human | |
|---|---|
| Location: | 12q13.3 |
| Sequence: | 12; NC_000012.12 (57516588..57520517, complement) |
Nucleus. Cytoplasm.




PMID: 29083303 by Samandi S., et al. Deep transcriptome annotation enables the discovery and functional characterization of cryptic small proteins.
*More publications can be found for each product on its corresponding product page