This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

2 Citations 16 Q&As



Facts about High affinity nerve growth factor receptor.

However, NTF3 only supports axonal extension through NTRK1 but has no effect on neuron survival (By similarity). Upon dimeric NGF ligand-binding, undergoes homodimerization, autophosphorylation and activation (PubMed:1281417).
| Human | |
|---|---|
| Gene Name: | NTRK1 |
| Uniprot: | P04629 |
| Entrez: | 4914 |

| Belongs to: |
|---|
| protein kinase superfamily |

DKFZp781I14186; EC 2.7.10; EC 2.7.10.1; MTChigh affinity nerve growth factor receptor; Neurotrophic tyrosine kinase receptor type 1; neurotrophic tyrosine kinase, receptor, type 1; NTRK1; NTRK-1; p140-TrkA; TRK1-transforming tyrosine kinase protein; TrkA; Trk-A; TRKAOncogene TRK; TRKTRK1; tyrosine kinase receptor A
Mass (kDA):
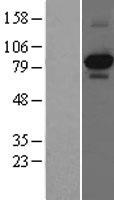
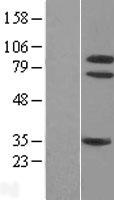
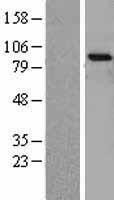
87.497 kDA

| Human | |
|---|---|
| Location: | 1q23.1 |
| Sequence: | 1; NC_000001.11 (156815750..156881850) |
Isoform TrkA-I is found in most non-neuronal tissues. Isoform TrkA-II is primarily expressed in neuronal cells. TrkA-III is specifically expressed by pluripotent neural stem and neural crest progenitors.
Cell membrane; Single-pass type I membrane protein. Early endosome membrane; Single-pass type I membrane protein. Late endosome membrane; Single-pass type I membrane protein. Recycling endosome membrane; Single-pass type I membrane protein. Rapidly internalized after NGF binding (PubMed:1281417). Internalized to endosomes upon binding of NGF or NTF3 and further transported to the cell body via a retrograde axonal transport. Localized at cell membrane and early endosomes before nerve growth factor (NGF) stimulation. Recruited to late endosomes after NGF stimulation. Colocalized with RAPGEF2 at




PMID: 2927393 by Martin-Zanca D., et al. Molecular and biochemical characterization of the human trk proto- oncogene.
PMID: 7823156 by Shelton D.L., et al. Human trks: molecular cloning, tissue distribution, and expression of extracellular domain immunoadhesins.
*More publications can be found for each product on its corresponding product page