This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

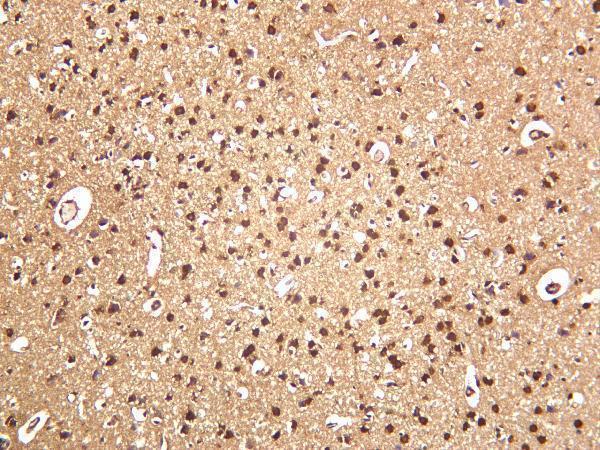
13 Citations 16 Q&As
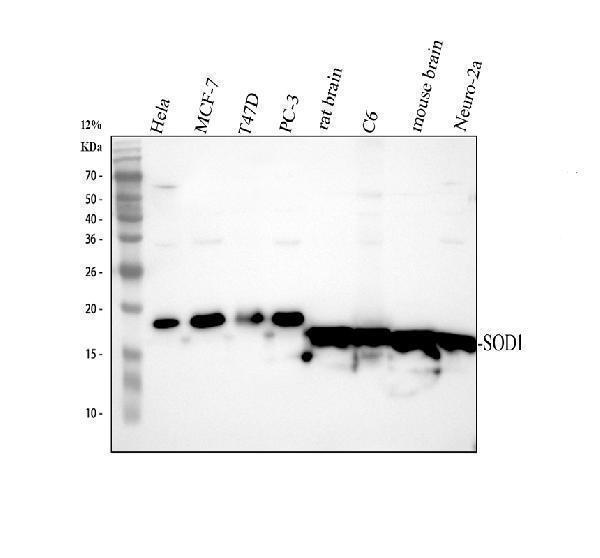
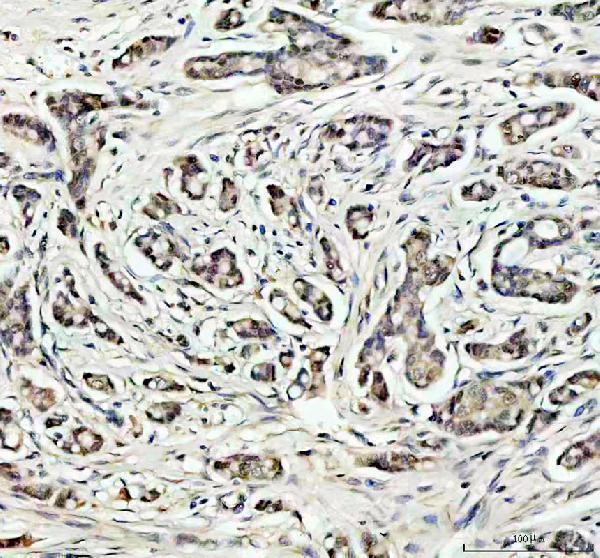
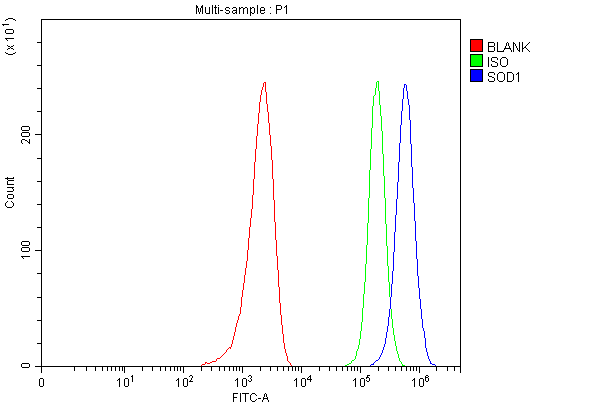
10 Citations 18 Q&As
1 Citations 5 Q&As
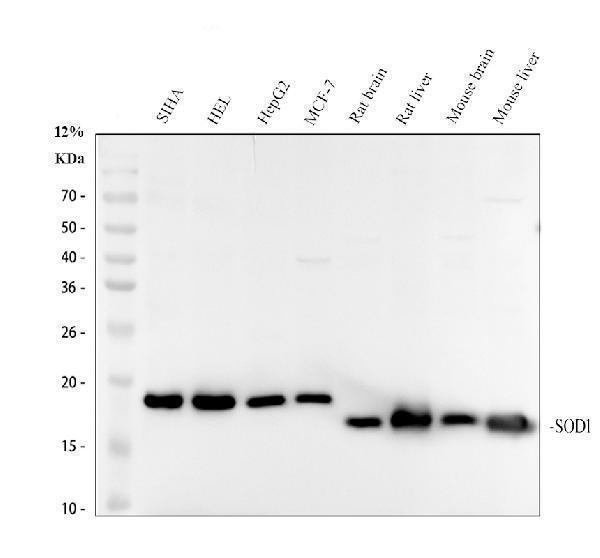
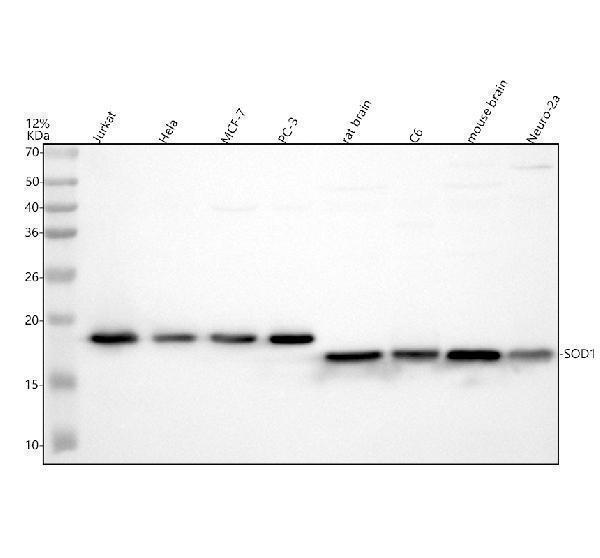
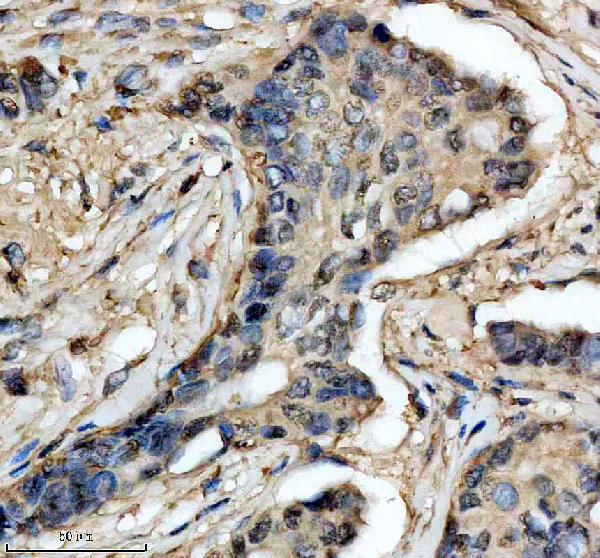


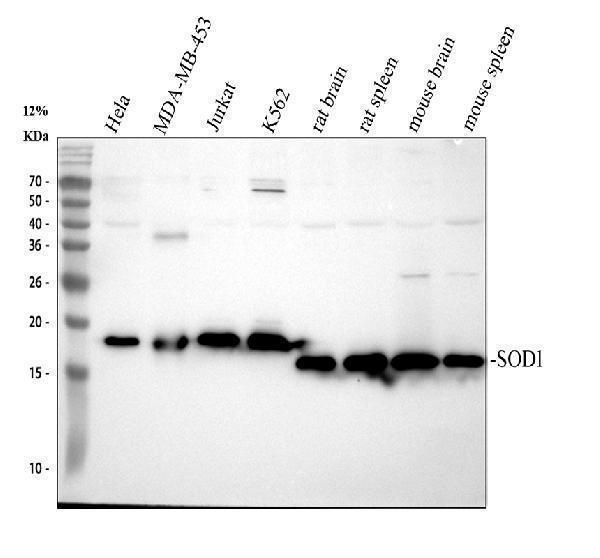
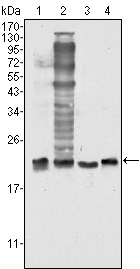
Facts about Superoxide dismutase [Cu-Zn].

| Human | |
|---|---|
| Gene Name: | SOD1 |
| Uniprot: | P00441 |
| Entrez: | 6647 |

| Belongs to: |
|---|
| Cu-Zn superoxide dismutase family |

ALS; ALS1; amyotrophic lateral sclerosis 1 (adult); Cu; Cu/Zn superoxide dismutase; CuZn SOD; Cu-Zn SOD; EC 1.15.1.1; homodimer; hSod1; indophenoloxidase A; Ipo1; IPOA; SOD; SOD, cytosolic; SOD, Soluble; SOD1; superoxide dismutase [Cu-Zn]; Superoxide dismutase 1; superoxide dismutase 1, soluble; Zn superoxide dismutase, EC 1.15.1.110superoxide dismutase, cystolic
Mass (kDA):
15.936 kDA

| Human | |
|---|---|
| Location: | 21q22.11 |
| Sequence: | 21; NC_000021.9 (31659693..31668931) |
Cytoplasm. Mitochondrion. Nucleus. Predominantly cytoplasmic; the pathogenic variants ALS1 Arg-86 and Ala-94 gradually aggregates and accumulates in mitochondria.





PMID: 6577438 by Sherman L., et al. Nucleotide sequence and expression of human chromosome 21-encoded superoxide dismutase mRNA.
PMID: 3160582 by Levanon D., et al. Architecture and anatomy of the chromosomal locus in human chromosome 21 encoding the Cu/Zn superoxide dismutase.
*More publications can be found for each product on its corresponding product page