This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

3 Citations 3 Q&As
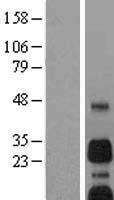
Facts about BCL2/adenovirus E1B 19 kDa protein-interacting protein 3.
Involved in mitochondrial quality control through its interaction with SPATA18/MIEAP: in response to mitochondrial damage, participates in mitochondrial protein catabolic process (also named MALM) resulting in the degradation of damaged proteins within mitochondria. The physical interaction of SPATA18/MIEAP, BNIP3 and BNIP3L/NIX in the mitochondrial outer membrane regulates the opening of a pore in the mitochondrial double membrane in order to mediate the translocation of lysosomal proteins in the cytoplasm to the mitochondrial matrix.
| Human | |
|---|---|
| Gene Name: | BNIP3 |
| Uniprot: | Q12983 |
| Entrez: | 664 |

| Belongs to: |
|---|
| NIP3 family |

BCL2/adenovirus E1B 19 kDa protein-interacting protein 3; BCL2/adenovirus E1B 19kDa interacting protein 3; BCL2/adenovirus E1B 19kD-interacting protein 3; BNIP3; Nip3
Mass (kDA):
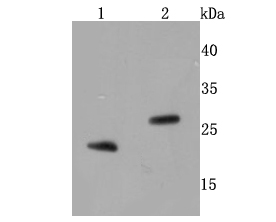
27.832 kDA

| Human | |
|---|---|
| Location: | 10q26.3 |
| Sequence: | 10; NC_000010.11 (131967683..131982013, complement) |
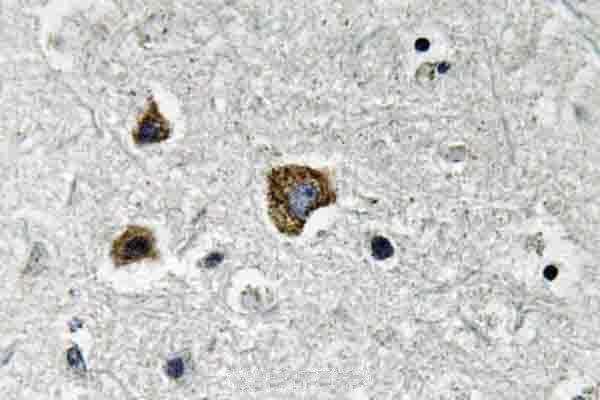
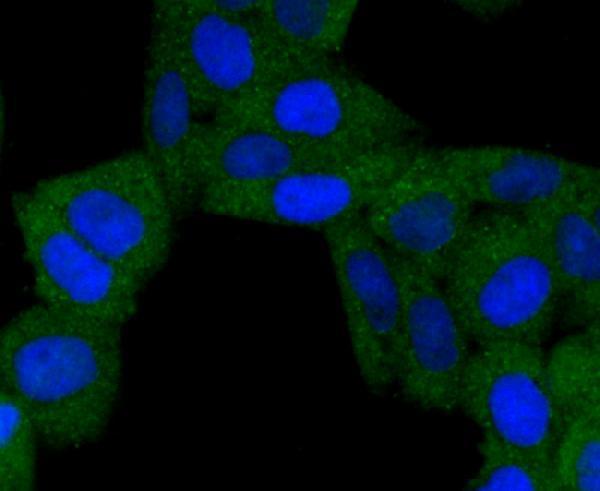
Mitochondrion. Mitochondrion outer membrane; Single-pass membrane protein. Coexpression with the EIB 19-kDa protein results in a shift in NIP3 localization pattern to the nuclear envelope. Colocalizes with ACAA2 in the mitochondria. Colocalizes with SPATA18 at the mitochondrion outer membrane.





PMID: 9396766 by Chen G., et al. The E1B 19K/Bcl-2-binding protein Nip3 is a dimeric mitochondrial protein that activates apoptosis.
PMID: 7954800 by Boyd J.M., et al. Adenovirus E1B 19 kDa and Bcl-2 proteins interact with a common set of cellular proteins.
*More publications can be found for each product on its corresponding product page