This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

1 Citations
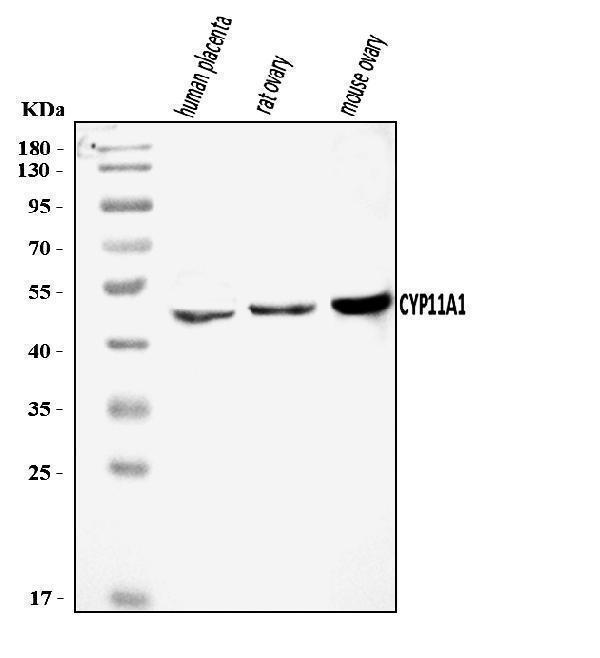
Facts about Cholesterol side-chain cleavage enzyme, mitochondrial.

| Human | |
|---|---|
| Gene Name: | CYP11A1 |
| Uniprot: | P05108 |
| Entrez: | 1583 |

| Belongs to: |
|---|
| cytochrome P450 family |

Cholesterol desmolase; cholesterol monooxygenase (side-chain-cleaving); cholesterol side-chain cleavage enzyme, mitochondrial; CYPXIA1; Cytochrome P450 11A1; Cytochrome P450(scc); cytochrome P450, family 11, subfamily A, polypeptide 1; cytochrome P450C11A1; EC 1.14.15; EC 1.14.15.6; steroid 20-22-lyase; subfamily XIA (cholesterol side chain cleavage)
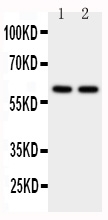
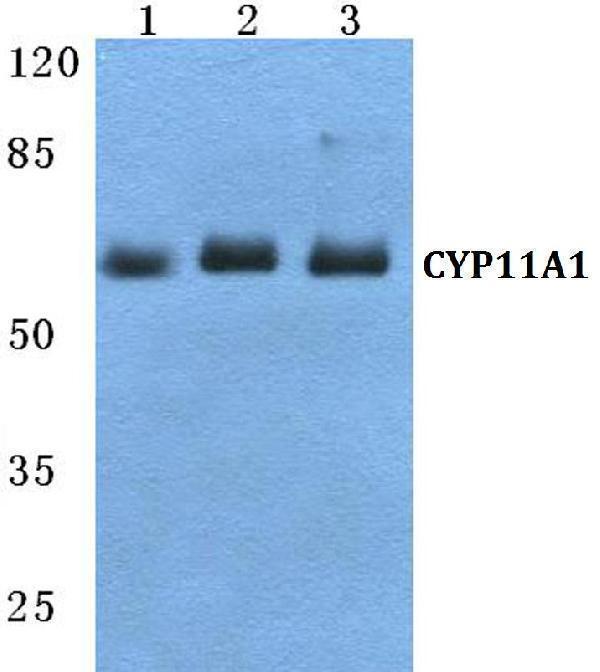
Mass (kDA):
60.102 kDA

| Human | |
|---|---|
| Location: | 15q24.1 |
| Sequence: | 15; NC_000015.10 (74337762..74367646, complement) |
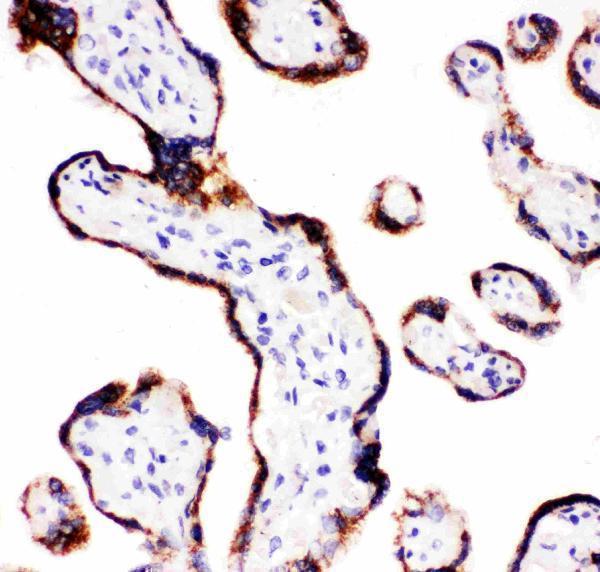
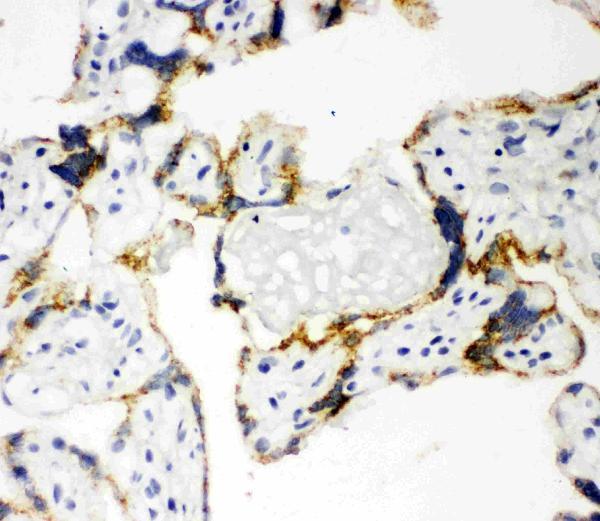
Mitochondrion inner membrane; Peripheral membrane protein. Localizes to the matrix side of the mitochondrion inner membrane.




PMID: 3024157 by Chung B.-C., et al. Human cholesterol side-chain cleavage enzyme, P450scc: cDNA cloning, assignment of the gene to chromosome 15, and expression in the placenta.
PMID: 3038854 by Morohashi K., et al. Gene structure of human cytochrome P-450(SCC), cholesterol desmolase.
*More publications can be found for each product on its corresponding product page