This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

1 Citations 4 Q&As



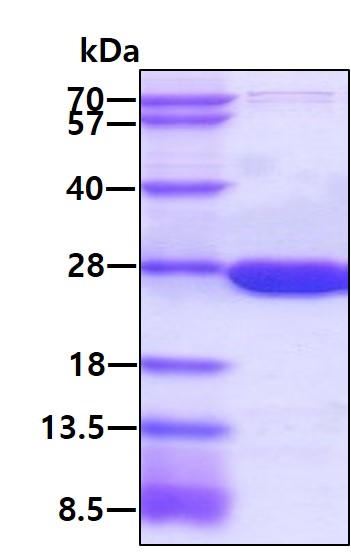
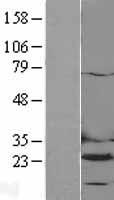
Facts about Dihydrofolate reductase.

Catalyzes an essential reaction for de novo glycine and purine synthesis, and for DNA precursor synthesis. Binds its own mRNA and that of DHFR2.
| Human | |
|---|---|
| Gene Name: | DHFR |
| Uniprot: | P00374 |
| Entrez: | 1719 |

| Belongs to: |
|---|
| dihydrofolate reductase family |

DHFR; DHFRP1; Dihydrofolate Reductase; DYR; EC 1.5.1.3
Mass (kDA):
21.453 kDA

| Human | |
|---|---|
| Location: | 5q14.1 |
| Sequence: | 5; NC_000005.10 (80626226..80654983, complement) |
Widely expressed in fetal and adult tissues, including throughout the fetal and adult brains and whole blood. Expression is higher in the adult brain than in the fetal brain.
Mitochondrion. Cytoplasm.




PMID: 6323448 by Chen M.-J., et al. The functional human dihydrofolate reductase gene.
PMID: 6687716 by Masters J.N., et al. The nucleotide sequence of the cDNA coding for the human dihydrofolic acid reductase.
*More publications can be found for each product on its corresponding product page