This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

5 Citations 5 Q&As
1 Citations 17 Q&As
5 Citations 16 Q&As
10 Citations 4 Q&As
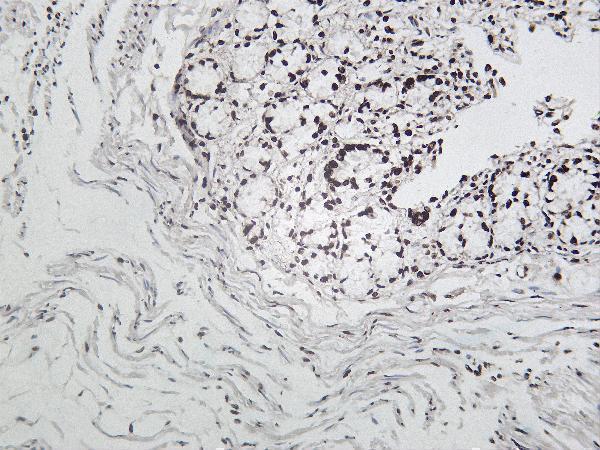
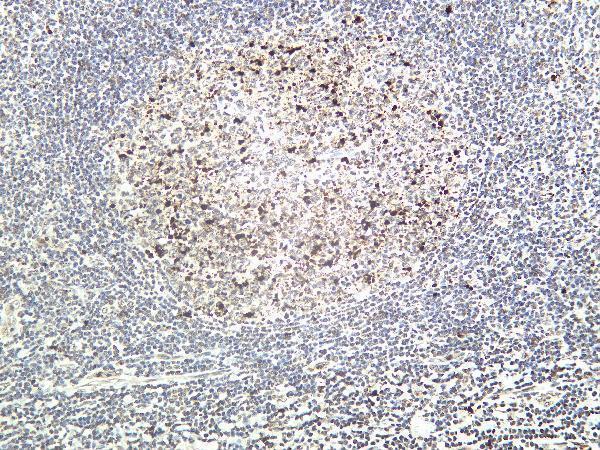
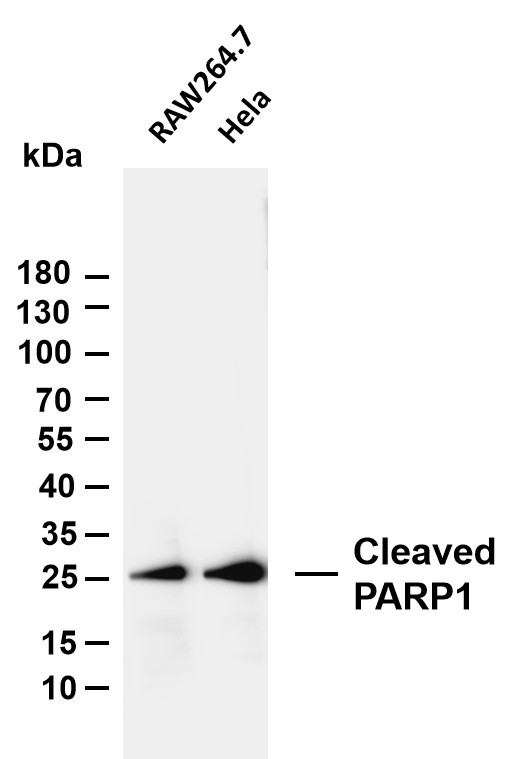
Facts about Poly [ADP-ribose] polymerase 1.

With EEF1A1 and TXK, forms a complex that serves as a T-helper 1 (Th1) cell-specific transcription factor and binds the promoter of IFN-gamma to directly regulate its own transcription, and is consequently involved importantly in Th1 cytokine production (PubMed:17177976). Required for PARP9 and DTX3L recruitment to DNA damage sites (PubMed:23230272).
| Human | |
|---|---|
| Gene Name: | PARP1 |
| Uniprot: | P09874 |
| Entrez: | 142 |

| Belongs to: |
|---|
| No superfamily |

ADP-ribosyltransferase (NAD+; poly (ADP-ribose) polymerase); ADPRT 1; ADPRT; ADPRTADP-ribosyltransferase NAD(+); EC 2.4.2; EC 2.4.2.30; NAD(+) ADP-ribosyltransferase 1; PARP apoptosis; PARP; PARP1; PARP-1; PARPADPRT1; poly (ADP-ribose) polymerase 1; poly (ADP-ribose) polymerase family, member 1; poly [ADP-ribose] polymerase 1; poly(ADP-ribose) polymerase; poly(ADP-ribose) synthetase; poly(ADP-ribosyl)transferase; Poly[ADP-ribose] synthase 1; PPOL; PPOLpADPRT-1
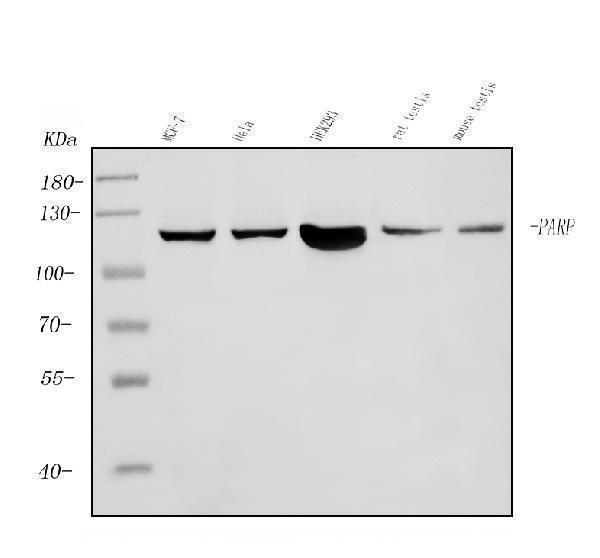
Mass (kDA):
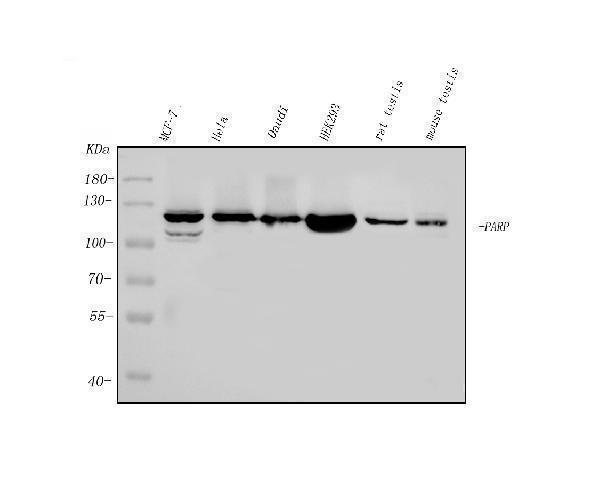
113.084 kDA

| Human | |
|---|---|
| Location: | 1q42.12 |
| Sequence: | 1; NC_000001.11 (226360691..226408093, complement) |
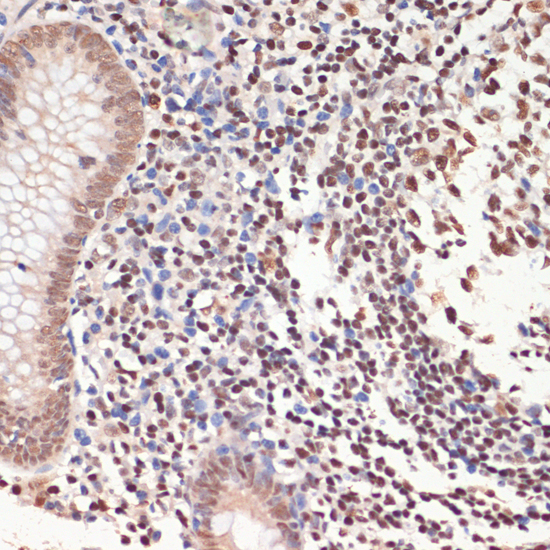
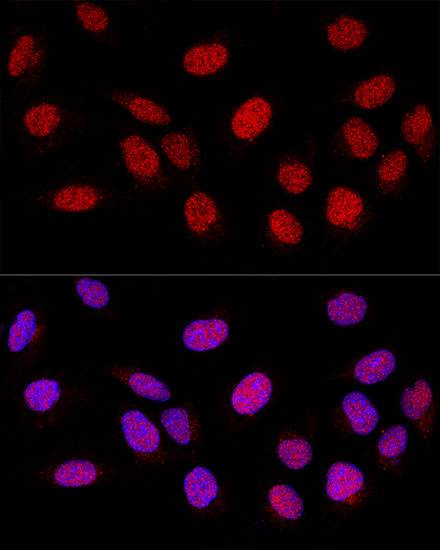
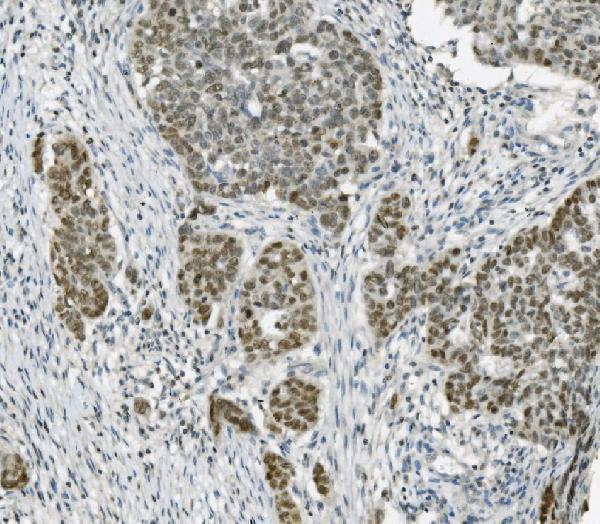
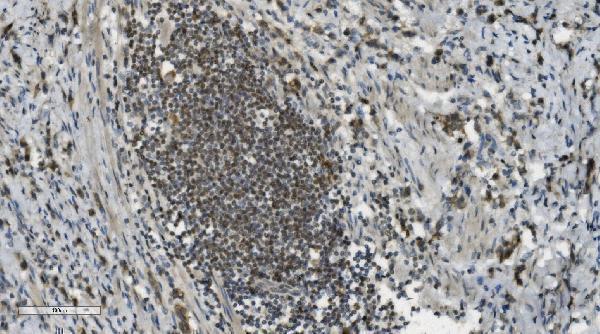
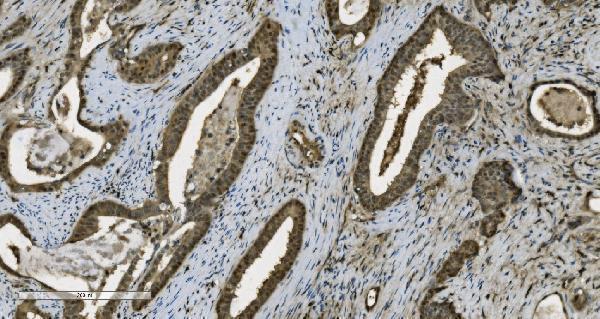
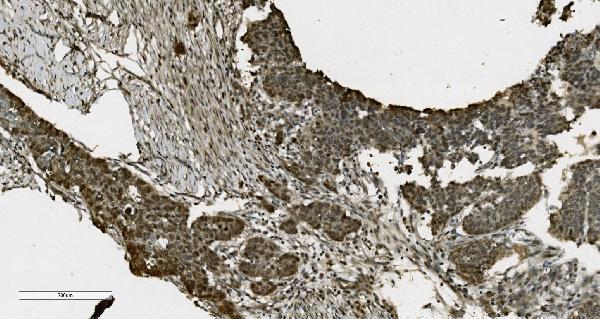
Nucleus. Nucleus, nucleolus. Chromosome. Localizes to sites of DNA damage.





PMID: 3120710 by Uchida K., et al. Nucleotide sequence of a full-length cDNA for human fibroblast poly(ADP-ribose) polymerase.
PMID: 2824474 by Kurosaki T., et al. Primary structure of human poly(ADP-ribose) synthetase as deduced from cDNA sequence.
*More publications can be found for each product on its corresponding product page