This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

2 Citations 6 Q&As
Facts about Doublesex- and mab-3-related transcription factor 1.

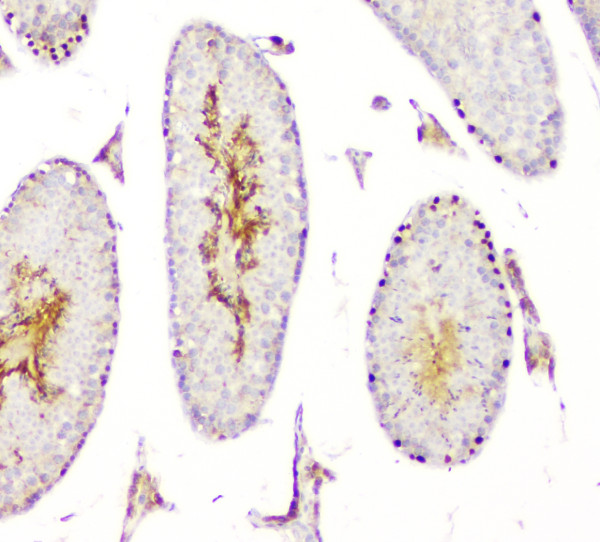
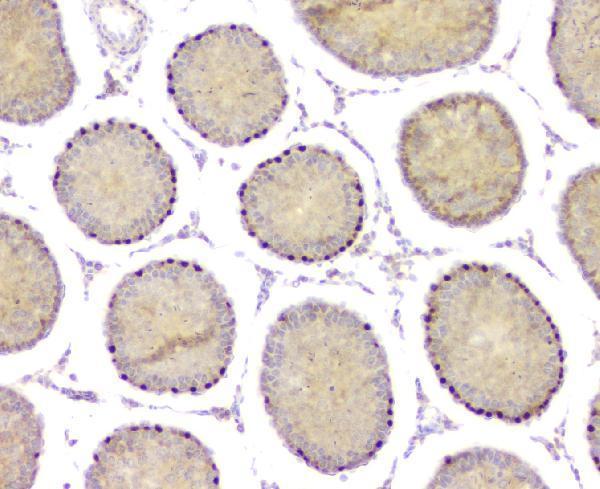
Acts both as a transcription repressor and activator: prevents meiosis by restricting retinoic acid (RA)-dependent transcription and repressing STRA8 expression and promotes spermatogonial growth by activating spermatogonial differentiation genes, such as SOHLH1. Also plays a key role in postnatal sex care by keeping testis determination and preventing feminization: represses transcription of feminine promoting genes such as FOXL2 and activates male-specific genes.
| Human | |
|---|---|
| Gene Name: | DMRT1 |
| Uniprot: | Q9Y5R6 |
| Entrez: | 1761 |

| Belongs to: |
|---|
| DMRT family |

DM domain expressed in testis 1; DMT1DM domain expressed in testis protein 1; doublesex and mab-3 related transcription factor 1; doublesex- and mab-3-related transcription factor 1
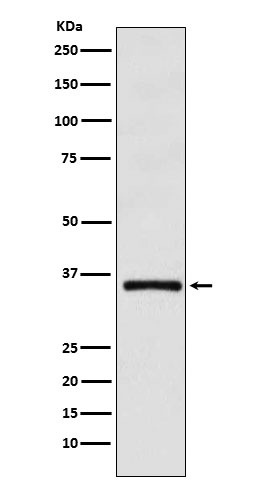
Mass (kDA):
39.473 kDA

| Human | |
|---|---|
| Location: | 9p24.3 |
| Sequence: | 9; NC_000009.12 (841647..969090) |
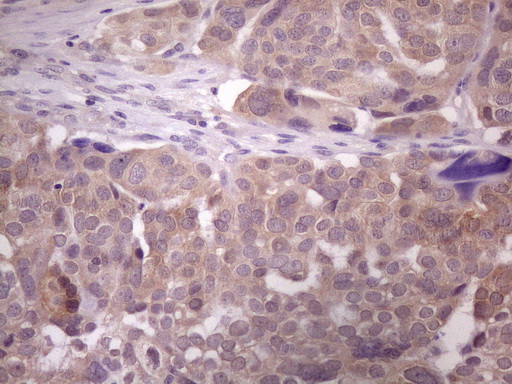
Testis-specific. Expressed in prostate cancer (at protein level).
Nucleus.



PMID: 10332030 by Raymond C.S., et al. A region of human chromosome 9p required for testis development contains two genes related to known sexual regulators.
PMID: 10857744 by Calvari V., et al. A new submicroscopic deletion that refines the 9p region for sex reversal.
*More publications can be found for each product on its corresponding product page