This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
A signaling cascade that performs an evolutionarily conserved function in organ size regulation by suppressing cell proliferation, inducing apoptosis, controlling stem/progenitor cell fates, and restricting cell size in specific conditions.
In developmental biology, the fundamental question is how an organ can stop growing after a particular size has reached. Organ growth is based on several cellular processes, including cell division and programmed cell death (or apoptosis). The Hippo signaling pathway contributes to restricting and promoting cell proliferation. As many cancers are marked by uncontrolled cell division, the signaling path for human cancer study has become increasingly important. The Hippo Pathway also plays a crucial role in stem cell and tissue-specific progenitor cell self-renovation and development.
It seems that the Hippo signals path is highly conserved. While most of the Hippo pathway components (Drosophila melanogaster) were identified in the fruit fly with mosaic genetic screening, ornithologists (genes associated to speciation events and therefore often retaining the same function in different species) were found in mammals. Drosophila, thus, helped to identify several genes that work as oncogenes or tumor suppressors in mammals.
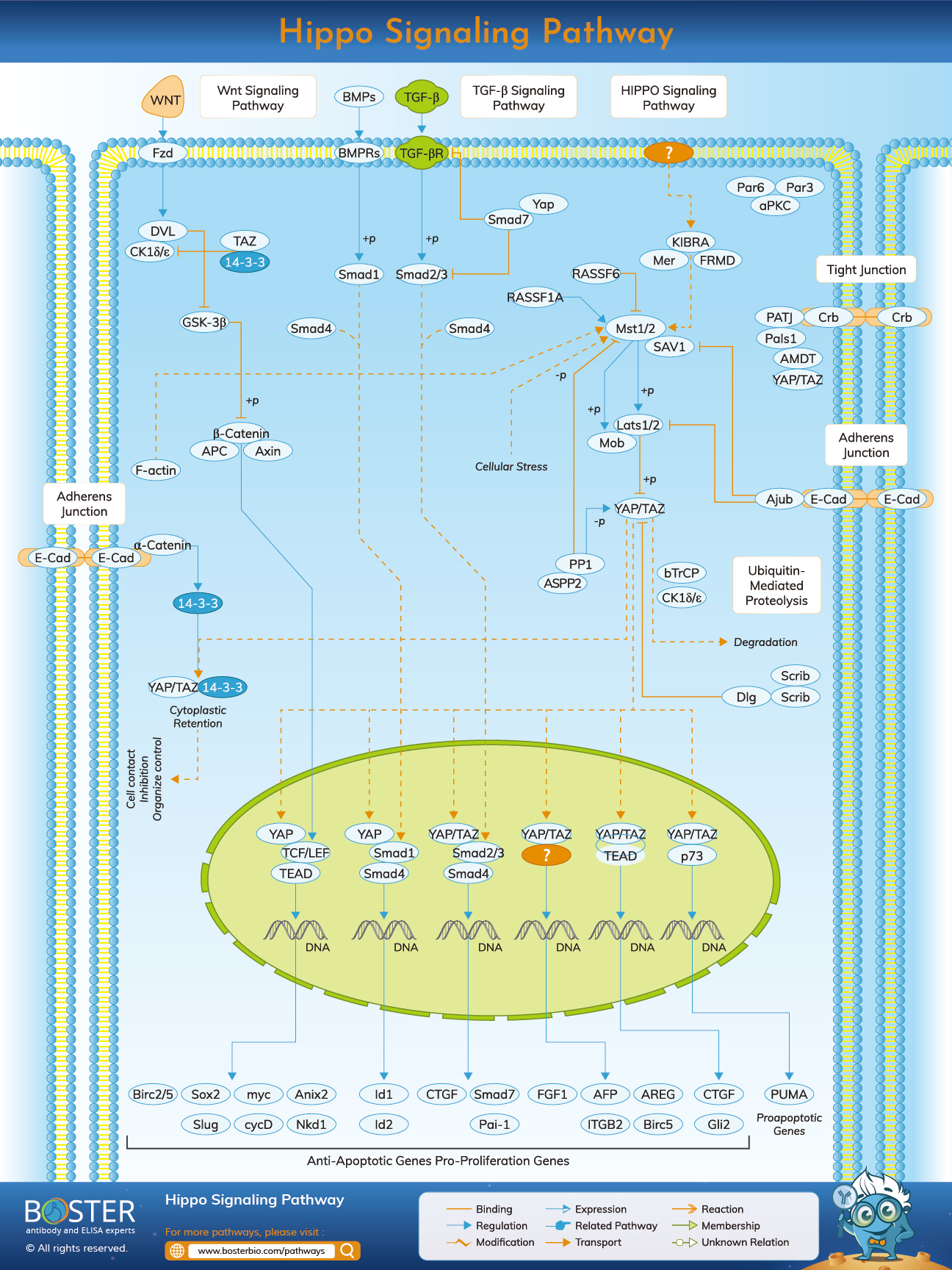
The Hippo signaling trajectories are a pathway that controls the organ sizes in animals via cell proliferation and apoptosis. Thus, it is also known as "the Salvador Warts Hippo (SWH) pathway." The route takes its name from one of the critical signals, the Hippo protein kinase (Hpo). This gene's mutations cause an overgrowth of tissues or a phenotype like a hippopotamus.
The Pathway, Hippo Signaling, is a crucial kinase cascade in which the protein kinase Warts is formed by Hpo phosphorylates (Drosophila) (Wts). A member of the Ste-20 family of protein kinases, Hpo (MST1/2 in mammals). This well-preserved serine/threonine kinase group controls many cell functions, including cell growth, death, and different stress reactions. Wts (LATS1/2 in mammals) is active as soon as it is phosphorylated. A malformed process of Hpo is carried out parallel with Msn, MAP4K4/6/7 (mammals), and Happy hour (mammals Happy, MAP4K1/2/3/5). Wts is a kinase associated with nuclear DBF-2. These kinases are recognized cell cycle advancement, growth, and development regulators. The activation of Wts is identified as two proteins: Salvador (Sav) and Mob as tumor suppressors (Mats). Sav (SAV1 in mammals) is a WW-domain protein that includes a sequence of amino acids that are highly conserved with tryptophan and an invariant proline. He may bind to Sav and its phosphorylate that works as a scaffold protein because this interactive Hpo-Sav supports Wts phosphorylation. Mats (MOBKL1A, B in mammals) that enable Mats to combine and enhance the kinase activity of Wts may also be phosphorylated and activated by Hpo.
The Wts may be activated, and the transcriptional co-activator Yorkie in-activated (Yki). Yki cannot bind DNA alone. Yki is connected to the Scalloped (Sd) transcription factor in its operational condition, and the Yki-Sd complex becomes located in the center. This enables many genes to be explicit, such as cyclin, which stimulates the development of the cell cycle, and diap1, which inhibits apoptosis, as the name implies. Yki also active bantam microRNA expression, a positive cell number growth regulator. Thus, Wts' inactivation of Yki limits the development of these growth regulators via transcriptional repression. Wts enhances the interaction of yki with proteins of 14-3-3 by phosphorylating yki at serine 168, helping to anchor Yki within the cytoplasm and preventing its transport to the nucleus. In mammals, two Yki orthologs are Yes-associated proteins (YAP) and PDZ-binding transcriptional co-activators (WWTR1, also known as TAZ). If enabled, YAP and TAZ may bind to many factors such as p73, Runx2, and various TEADs. In mouse and human epithelial cells, YAP controls Hoxa1 and Hoxc13 in vivo and in vitro
Including transmembrane protein Fat and other membrane-related proteins, the upstream regulators for cores Hpo/Wts kinase cascade. Fat (FAT1-4 in animals) may act as an atypical cadherin, although an extracellular ligand hasn't been found positively. While fat is known to link with another atypical cadherin, Dachsous (Ds), the function of Ds in tissue development is unknown. However, Fat is known as the upstream Hpo pathway regulator. Fat stimulates Hpo using a spread of the apical protein (Ex, FRMD6/Willin for mammals). The Kibra (KIBRA in Mammals) and the Merlin (Mer; NF2 in Mammals) interacts with other apically localized proteins, which are forming a complex called the Kibra-Ex-Mer (KEM). Ex and Mer constitute FERM domain proteins, whereas Kibra, like Sav, is a WW domain protein. The KEM Complex interacts physically with the Hpo kinase cascade, locating the core kinase cascade for activation on the plasma membrane. The suppression of unconventional Dachs of myosin may also control Wts regardless of Ex/Hpo. Dachs may normally bind and enhance Wts degradation.
The Hippo signaling route is comprised of two YAP/YAZ variants. One type is phosphorylated YAP/TAZ, directly triggered by LATS1/2 kinase phosphorylation. In turn, many serine YAP and TAZ residues are phosphorylated and may lead to a 14-3-3 or proteasomal cytoplasm retention of YAP/TAZ. The hypophosphorylated YAP/TAZ is another kind of TAP/YAZ. The hypophosphorylated YAP/TAZ module translocates into the nuclear and induces relative gene expression when the Hippo signaling pathways' kinase module is inactivated.
In mediating the Wnt signaling route, β-catenin as a regulator plays a crucial function. YAP/TAZ retain cytoplasmic β-catenin; β-trCP recruits for destruction complex in certain situations. This alteration causes downregulation of nuclear β-catenin and subsequently suppresses Wnt. Cytoplasmic YAP and TAZ also adjust the WNT signal by holding β-catenin and scalpel (DVL). SHP2 tyrosine phosphatase, YAP, and TAZ transmit the nucleus from the cytoplasm to the nucleus. It stimulates β-catenin-dependent transcription when it is situated in nuclear. The interaction of SHP2 with the phosphorylated YAP and TAZ translates into the cytoplasm. YAP and TAZ monitor SHP2 and its role for nuclear and cytoplasmic localization.
The TGFβ/BMP binding to its serine-threonine kinase receptor leads to phosphorylation and activation of SMAD. The cytoplasmic YAP and TAZ sequester SMAD2/3 in the cytoplasm and subsequently block the signaling mediated by the SMAD2/3 route, as seen in the picture above in Hippo's image. Nuclear YAP enables transcription mediated by SMAD1. When TgFβ is activated, a complex, TEA domain family (TEAD) of nuclear YAP/TAZ and SMAD2/3 transcribers will be formed, and the protumorigenic transcriptional program will coordinate. YAP and TAZ also encourage transcription of many receptors and ligands to mediate the crosstalk and possible feedback loops of endocrine and paracrine signals.