This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
A conserved pathway that regulates crucial aspects of cell fate determination, cell migration, cell polarity, neural patterning and organogenesis during embryonic development.
The discovery of Wnt signaling was influenced by research on oncogenic (cancer-causing) retroviruses. In 1982, Roel Nusse and Harold Varmus infected mice with mouse mammary tumor virus in order to mutate mouse genes to see which mutated genes could cause breast tumors. They identified a new mouse proto-oncogene that they named int1 (integration 1)
Continued research led to the discovery of further int1-related genes; however, because those genes were not identified in the same manner as int1, the int gene nomenclature was inadequate. Thus, the int/Wingless family became the Wnt family and int1 became Wnt1. The name Wnt is a portmanteau of int and Wg and stands for "Wingless-related integration site".
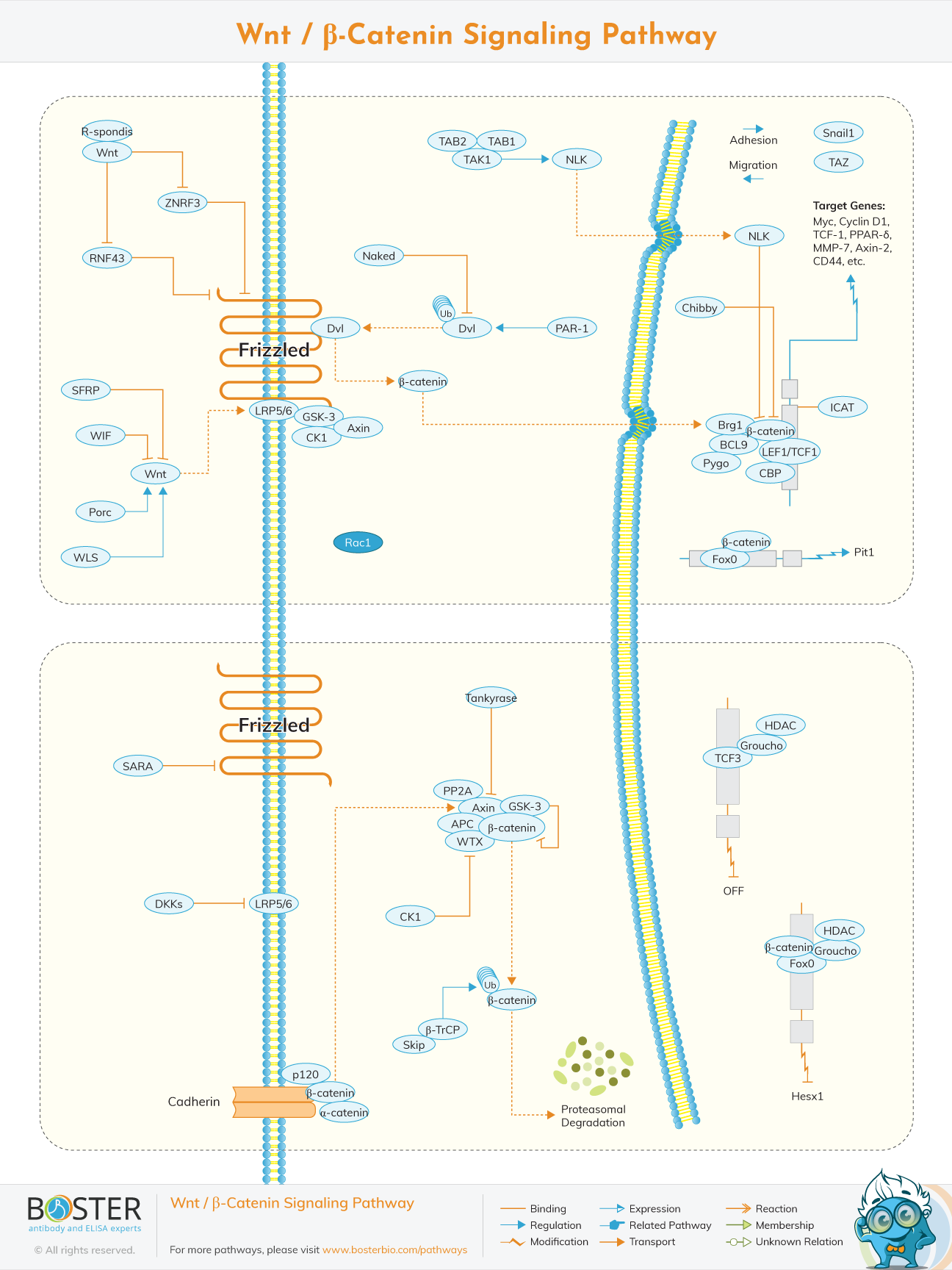
Wnt proteins are a family of cysteine-rich glycoproteins that play a critical role during development and in cancer. Our Wnt signaling pathway highlights the three Wnt signaling pathways: the canonical beta-catenin and the non-canonical planar cell polarity, and Wnt/Ca2+ pathways.
Wnt proteins regulate the proliferation of cells. In comparison to other growth factors, Wnt signals have several unique properties, including a short range of action. Thereby, Wnts predominantly mediate signaling locally, between neighboring cells.
Stabilized beta-catenin enters the nucleus and associates with T cell factor (TCF)/lymphoid enhancer factor (LEF) transcription factors, which leads to the transcription of Wnt target genes such as cyclin D1, PPARD, and twin.
In the absence of a Wnt signal, TCF/LEF family members interact with transcriptional inhibitors such as Groucho, which serve to repress Wnt signaling. The repressing effect of Groucho is mediated by interactions with histone deacetylases (HDAC), which are thought to make DNA refractory to transcriptional activation.
Negative regulators of beta-catenin signaling include ICAT and duplin, which directly bind to beta-catenin preventing it from interacting with TCF/LEF7. In addition to its role in Wnt signaling, beta-catenin plays a role in cell adhesion through binding to the cytoplasmic domain of type 1 cadherins and linking them through alpha-catenin to the actin cytoskeleton.
On a molecular level, these different biological activities are mediated through different intracellular signaling pathways. Axis duplication and cell transformation are based on the activation of the canonical Wnt pathway that involves the multifunctional protein β-catenin. In the absence of Wnt, β-catenin is targeted to a multimeric protein complex called destruction complex for its phosphorylation.
The coordinated alignment of cell polarity across the tissue plane is referred to as planar cell polarity (PCP). Asymmetric division of cortical PCP components and intercellular communication to coordinate polarity between nearby cells are critical to the formation of PCP.
Wnt signaling via Frizzled receptors enables asymmetric cytoskeletal structure and cell polarization by causing actin cytoskeleton changes in the planar cell polarity pathway. Dsh initiates two separate pathways that result in the activation of the small GTPases Rho and Rac. Rho activation necessitates the activation of Daam-1, which in turn activates the Rho-associated kinase ROCK. Rac activation occurs independently of Daam-1 and promotes Jun Kinase (JNK) activity.
The creation of PCP, like many other forms of cell polarity, includes a global orienting cue, asymmetric segregation of specialised polarity proteins, and translation of polarity information into polarized outputs. However, unlike other kinds of cell polarity, the PCP processes we presently understand entail cell-to-cell interaction, allowing for polarity alignment across long distances.
Katharina Nübler-Jung used the term "planar polarity" to describe the structured polarization of cells in the plane of an epithelium in the 1980s. Nübler-Jung and other researchers conducted investigations on the cuticles of numerous insects in the prior decades, which were both accessible to experimental manipulation and demonstrated clear cellular polarity as evidenced by the ordered orientation of hairs, bristles or scales. The phrase ‘tissue polarity' has largely fallen out of usage, while the phrase ‘planar cell polarity', which may be reduced to PCP, has been warmly adopted by fans of three letter acronyms.
Wnt signaling via Frizzled receptors can also result in intracellular calcium release. Knypek and Ror2 are frizzled co-receptors associated in this pathway. Other intracellular second messengers involved in this route are heterotrimeric G-proteins, phospholipase C (PLC), and protein kinase C. (PKC).
The particular genes activated by the Wnt/Ca2+ pathway are unclear, however NFAT, a transcription factor controlled by the calcium/calmodulin-dependent protein phosphatase calcineurin, appears to be essential. The Wnt/Ca2+ signaling pathway is vital for cell attachment and motility during gastrulation.
The Wnt/Ca(2+) signaling cascade is less well understood than its canonical cousin, the Wnt/-catenin pathway. Non-canonical Wnt signaling pathways include the planer cell polarity route, the Wnt-RAP1 signaling system, the Wnt-Ror2 signaling route, the Wnt-PKA pathway, the Wnt-GSK3MT route, the Wnt-aPKC pathway, the Wnt-RYK pathway, the Wnt-mTOR pathway, and the Wnt/calcium signaling pathway.
WNT signaling mutations have also been linked to oral disorders. Loss-of-function mutations in the WIF1 WNT inhibitor gene have been linked to salivary gland oncogenesis. Gardner syndrome, which includes multiple jaw and tooth cysts as well as intestinal polyposis, is also caused by APC gene mutations. AXIN2 mutations cause tooth agenesis (the lack of one or more teeth during development), with nonsense AXIN2 mutations causing severe permanent tooth agenesis through activating the WNT signaling pathway.
A nonsense mutation in WNT10A is responsible for severe hypodontia in odonto-onycho-dermal dysplasia, a rare ectodermal dysplasia characterized by keratoderma, dry hair and hyperkeratosis of the skin . Mutations in the human PORCN (porcupine) gene, which is a regulator of WNT signaling, cause focal skin hypoplasia, which is characterized by extensive dermal hypoplasia. WNT5A is increased in psoriasis whereas WNT Inhibitory Factor (WIF) is downregulated and pathway analysis suggested that this might be due to activation of non-canonical pathways.
The idea that WNT is important for bone development gained traction in the early 2000s, when several studies found that an inactivating mutation in LRP5 was linked to osteoporosis-pseudoglioma syndrome (OPPG), which is characterized by lower bone mass and strength and a higher risk of bone fractures. hese findings highlighted the importance of LRP5 for bone development and pioneered the interest in WNT’s role in bone development and disease.
A notable feature of the liver, unlike other tissues, is its ability to regenerate post-injury, driven by the WNT/β-catenin signaling pathway. Loss of β-catenin and other components of the WNT signaling pathway causes delayed liver regeneration following a partial hepatectomy. β-Catenin is upregulated in fibrotic human liver tissue, and increased β-catenin levels in rat hepatic stellate cells were associated with collagen production and proliferation
WNT4-mediated activation of β-catenin at different intervals after acute kidney injury results in recovery from renal injury by promoting cell cycle progression. WNT7B derived from macrophages activated WNT signaling to confer protection and trigger the repair process after ischemia reperfusion injury in mice. β-Catenin signaling was also reported to reduce Bax-mediated apoptosis and improve cell survival after induction of metabolic stress in proximal tubular epithelial cells.
Activation of canonical WNT signaling enhanced MMP-2, MMP-3, MMP-7, MMP-9, and MT3-MMP expression in mice, indicating that WNT modulates MMP production and consequently lung inflammation. Increased levels of matrix metalloproteinases (MMPs) and pro-inflammatory cytokines such as IL1, IL6, IL8 and IL15 are general features of lung inflammation. The canonical WNT signaling pathway has been shown to regulate the expression of both MMPs and pro-inflammatory cytokines.