This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
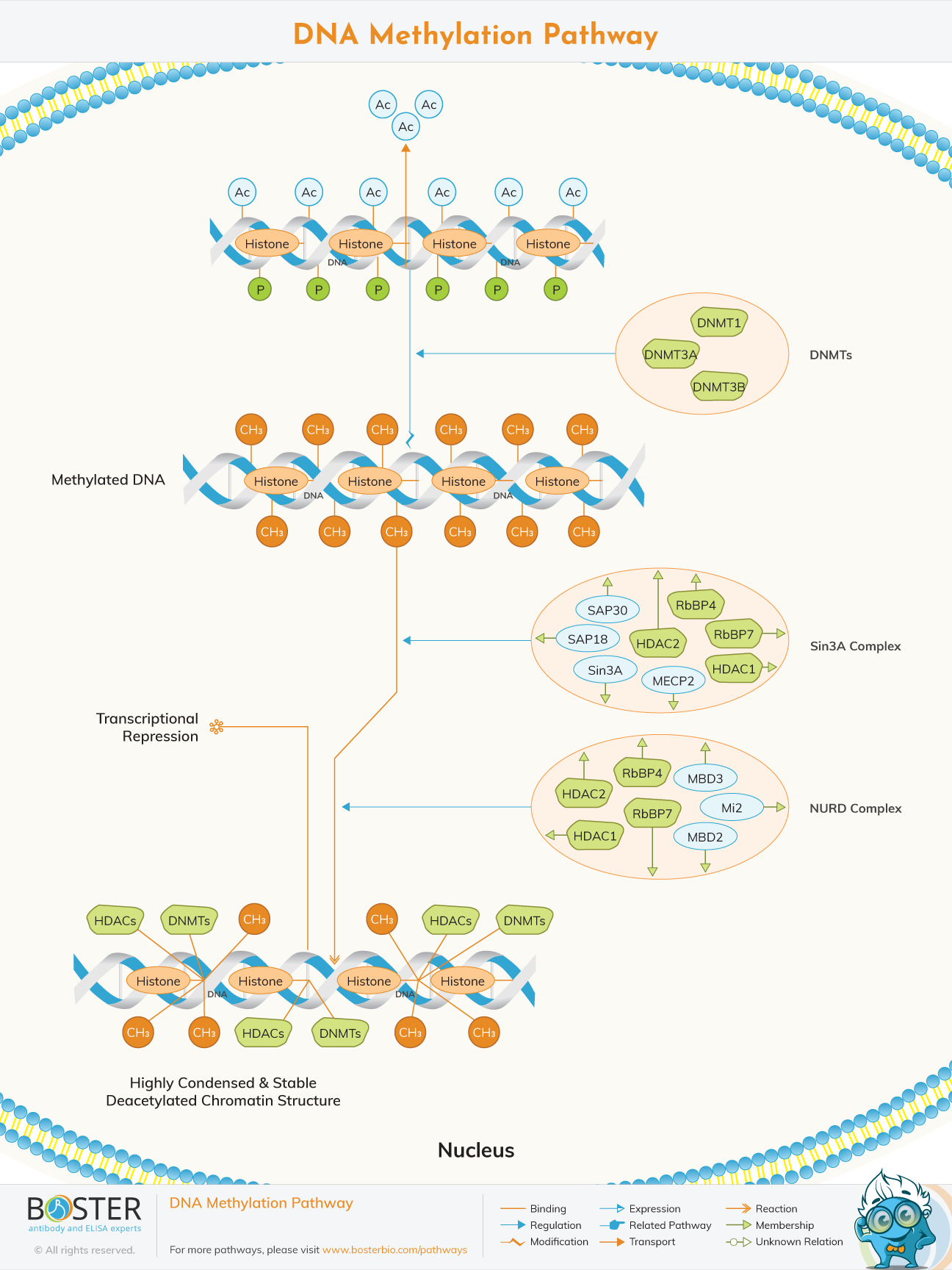
DNA methylation is an epigenetic modification which plays an important role in chromatin organization and gene expression. DNA methylation can silence genes and repetitive elements through a process which leads to the alteration of chromatin structure.
Transcription repression is an important mechanism for precise control of gene expression. Transcription repressors are directly associated with their target genes through DNA binding domains or indirectly through interactions with other DNA binding proteins. In order to selectively inhibit transcription, the repressor protein can mask the transcription activation domain, block the interaction of the activator with other components of the transcription mechanism, or replace the activator from the DNA. In addition, DNA response elements can exert an allosteric effect on transcription regulators, so that the regulator can activate transcription in the context of one gene, but repress transcription in another gene.
DNA methylation is a covalent modification in which the 50 position of cytosine is methylated in a reaction catalyzed by DNA methyltransferase (DNMT), and methionine is used as the methyl donor. In mammals, this modification occurs in CpG dinucleotides and can be catalyzed by three different enzymes, DNMT1, DMNT3a, and DNMT3b.
When the number of methylation sites increases and reaches a threshold that causes gene silencing to spread in DNA fibers, the inhibition mechanism is significantly different. Hypomethylation leads to chromosomal instability and may increase the expression of some proto-oncogenes. CpG island methylation can silence tumor suppressor genes and increase the possibility of mutations, which often occur in these regions.
DNA methylation interferes with transcription by preventing the basic transcription mechanism or the binding of ubiquitous transcription factors (TF), which require contact with cytosine (C) in the main groove of the double helix. The transcriptionally active chromatin is mainly unmethylated and has a high level of acetylated histone tails. Most mammalian TFs have GCrich binding sites, and many have CpG in their DNA recognition elements. CpG methylation prevents the binding of several of these factors. The methylation in CpG dinucleotides is carried out by one of the three known human DNA methyltransferases (DNMT1, DNMT3a, and DNMT3b), resulting in DNA with a high level of CpG methylation, but still mainly contains Acetylation of histone tails.
CpG methylation induces histone deacetylation, chromatin remodeling, and gene silencing through a transcription repressive complex, which includes SMRT (silencing mediators of retinoic acid and thyroid receptor), mSin3a, RbAp46/48, and the formation around mSin3a The two histone deacetylases HDAC1 and HDAC2.
The sequences within the genome can be divided into two different groups: CpG-poor regions and CpG islands. CpG islands are defined as having more than 500 bp and a GC content of more than 55% with an observed CpG/expected CpG ratio of 0.65. CpG islands are usually (but not always) found in the promoter region, and approximately 40% of genes contain CpG islands located at the end of 50 regions (promoter, untranslated region, and exon1).
The Nucleosome Remodelling and Deacetylase (NuRD) complex is one of the major chromatins remodelling complex found in cells. It plays a vital role in regulating gene transcription, progression of cell cycle and genome integrity. It is assembled as a result of DNA methylation and contains at least 6 proteins. They include;
• Enzymatic components, histone deacetylases HDAC 1 and HDAC 2 that remove acetyl groups from lysine residues on histone tails.
• Non-enzymatic components, methyl cytosine phosphate guanine binding domain proteins which are MBD 2 and MBD 3. MBD 2 binds cytosine methylated DNA while MBD 3 lacks this capacity to bind with high affinity.
• Histone binding proteins RbBP7 and RbBP4 that bind to Histone H4 and are thought to function as scaffolding proteins or chaperones.
• Chromodomain-helicase-DNA-binding proteins called Mi 2. They are of two types; Mi2- alpha and Mi2-beta. They both use ATP to reposition nucleosomes on DNA.
The Sin 3 complex gets it's name from the fact that it contains Sin 3A protein among many others. The Sin 3A protein is a paired amphipathic helix proteins that is found in humans and it's encoded by the SIN 3A gene. The complex has transcription corepressor activity. This complex has some components that's are unique to it and others can also be found in NuRD complex. The components found in both complexes are HDAC 1, HDAC 2, RbBP7 and RbBP4. The components unique to this complex include;
• The Sin 3A protein binds DNA which aids the complex in binding on the template DNA strand in the cell.
• SAP 18 and SAP 30 are proteins that mediate interactions with different polypeptides providing specificity of Sin 3 complexes.
Poor DNA methylation can lead to cancer due to lack of transcription repression which can lead to expression of oncogenes. DNA methylation has been concluded to be the cause of cardiovascular diseases if the methylation is too high. These diseases including atherosclerosis and heart failure for instance ischemic heart failure DNA methylation changes have been linked to change in gene expression causing changes in heart metabolism..