This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
ESCs (embryonic stem cells) are pluripotent cells derived from the ICM (inner cell mass) of blastocyst-stage embryos.
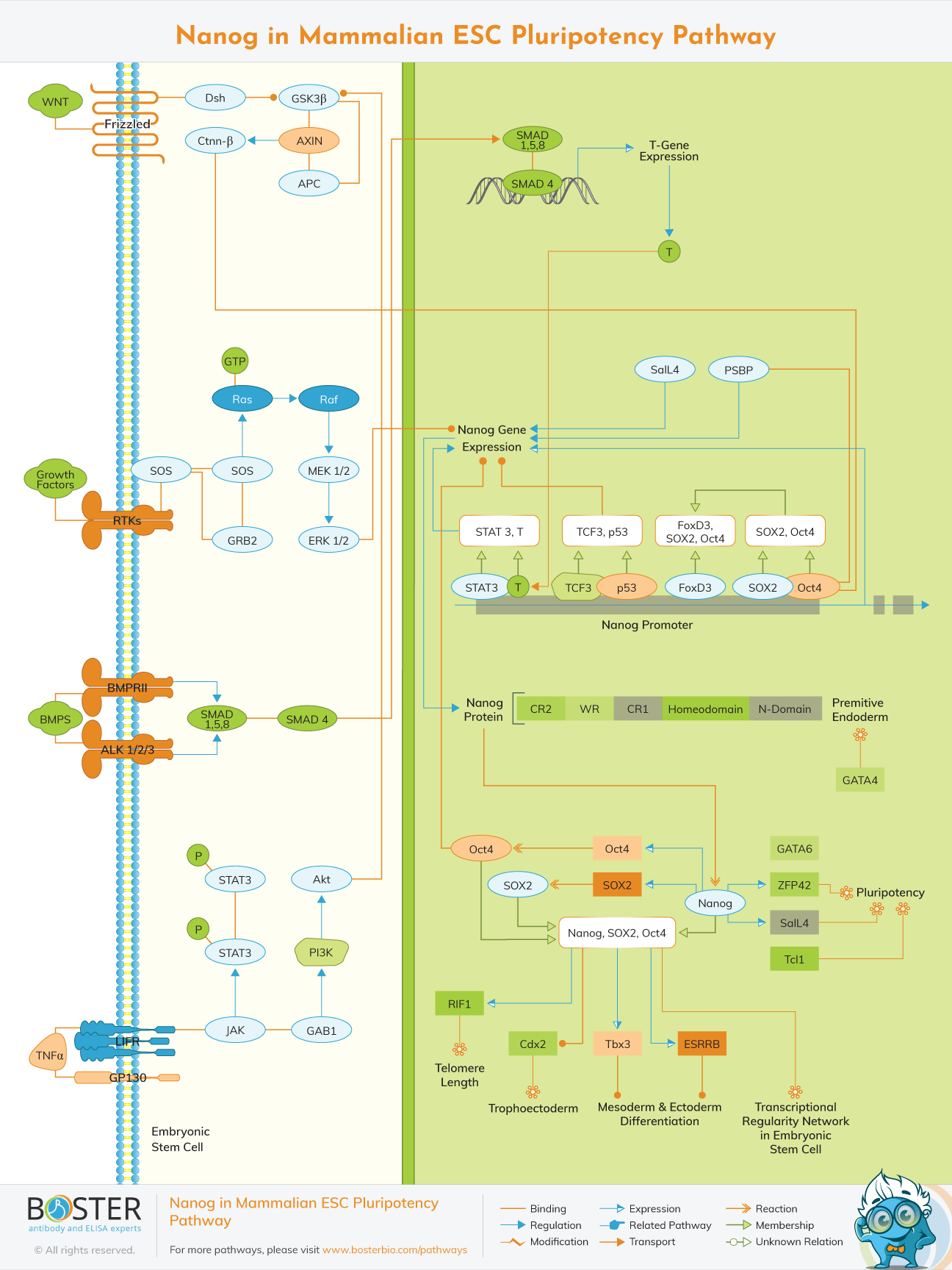
The transcription factor Nanog is a homeodomain-bearing protein that is transcribed specifically and monkey and human ESCs. Nanog protein consists of three domains with the homeodomain separating the N-terminal serine-rich 96 amino acids from the C-terminal 150 amino acids. Both N- and C-terminal domains in mouse Nanog show trans-activator function, but the activity of the C-terminal domain is much higher than the N-terminal domain. In human Nanog, transactivation potential is retained in the C-terminal domain but not in the N-terminal domain. The prominent feature of the C-terminal domain is the presence of a 10 pentapeptide repeat (WR), each starting with a tryptophan (W). This repeat is well conserved between humans and mice, although one of the tryptophans is replaced by a glutamine (Q) in human Nanog. In addition to WR, the C-terminal domain also contains another trans-activator module, the 58 amino acids region C-terminal to the WR (CD2). Both WR and CD2 can separately transactivate tthe expression of a reporter gene.
Nanog is a self-renewal and pluripotency-promoting transcription factor. It is the main regulator, capable of proliferating ESCs in the absence of specific external stimuli and inducing pluripotency in differentiated cells. LIF, BMP, and WNT activate ESCs via the JAK-STAT, SMAD, and beta-catenin pathways, resulting in Nanog expression and pluripotency maintenance.
Nanog expression is limited to pluripotent cells and is downregulated upon differentiation, although very little is understood about how it is regulated. A regulatory region 332 bp upstream of the transcriptional start site controls the transcription of the Nanog gene.
Oct4 and SOX2 are the main transcription factors that stimulate Nanog transcription in vitro and in vivo by binding to the Nanog promoter. High amounts of Nanog, on the other hand, promote ES cell self-renewal, whereas overexpressed Oct4 causes differentiation.
Nanog also plays a role in sustaining ESC pluripotency as a direct downstream effector of the LIF-STAT3 (Signal Transducer and Activator of Transcription-3) pathway. Nanog expression is also controlled by the BMP (Bone Morphogenetic Protein) pathway.
Low doses of BMPs stimulate mesoderm development of mouse ESCs by upregulating the mesoderm marker-T (Brachyury) and inhibit neural-ectoderm differentiation in the absence of LIF.In the presence of LIF, STAT3 interacts with Brachyury and binds the Nanog promoter, resulting in upregulation of Nanog expression. STAT3 is stimulated by JAK (Janus tyrosine Kinase), which is activated by the LIFR–GP130 (GP130 Transducer Chain) heterodimer on binding LIF.
BMPs belong to the TGF-Beta (Transforming Growth Factor-Beta) Superfamily and mediate signaling through their downstream effectors – SMAD1 [Sma and MAD (Mothers Against Decapentaplegic) Related Protein-1] or the closely related SMAD5 [Sma and MAD (Mothers Against Decapentaplegic) Related Protein-5] and SMAD8 (Sma and MAD (Mothers Against Decapentaplegic) Related Protein-8).
Nanog can directly interact with SMAD1 and prevent the coactivators from being recruited to active SMAD1 complexes, suppressing BMP signaling, limiting mesoderm development, and ultimately retaining the undifferentiated state of Mouse ESCs.
Catenin-Beta, a key regulator of WNT Pathway, also upregulates Nanog. In general, Catenin-Beta stability is negatively regulated through its phosphorylation by GSK3Beta (Glycogen Synthase Kinase-3-Beta), APC (Adenomatous Polyposis Coli), and AXIN (Axis Inhibitor) complex. Extracellular WNT proteins activate the canonical WNT signaling pathway by binding the Fz (Frizzled) seven transmembrane span receptor and stabilizing intracellular Ctnn-Beta, which then translocates to the nucleus, where it interacts with TCF (T-cell Factor)/ LEF (Lymphocyte Enhancer Factor) transcription factors to regulate expression of target genes. Catenin- Beta can also be stabilized by LIF Pathway. LIF stimulation leads to inactivation of GSK3Beta through the PI3K (Phosphoinositide 3-Kinase)/Akt pathway, thus stabilizing Ctnn-Beta.
It is involved in upregulation of Nanog through binding with Oct3/4 protein in the nucleus. Oct4 can also bind to a novel PSBP (pluripotential cell-specific SOX element-binding protein) leading to Nanog gene expression. Oct4 dominantly bound to the Octamer element, while PSBP preferentially bound to the SOX element. SalL4 [Sal-Like-4 (Drosophila)], a member of the Spalt-like protein family, interacts with Nanog and exists as a complex with Nanog. The SalL4 and Nanog complex resembles the network configuration for the Oct4 and SOX2 complex. Nanog and SalL4 co-target many genomic sites. SalL4 and Nanog also bind to and regulate the respective regulatory regions of their own genes.
Nanog's mechanism for regulating stem cell pluripotency isn't fully understood. Nanog may have a dual function, repressing genes involved in differentiation while activating those involved in self-renewal. Nanog binds to the promoter of REX1/ZFP42 (Zinc Finger Protein-42) and so controls its expression.Nanog's C-terminal transactivator is most likely responsible for this control. In controlling REX1 activity, SOX2, not Oct3/4, collaborates with Nanog. Nanog regulates endoderm transcription factors GATA4 (GATA Binding Protein-4) and GATA6 (GATA Binding Protein-6) as well. Nanog inhibits ESC differentiation by directly repressing GATA6, which in turn represses GATA4.
Nanog also controls the expression of the Oct4 and SOX2 genes, both of which are involved in pluripotency. Nanog also activates activities in conjunction with other elements to create ESC identity, such as Oct4 and SOX2. These factors control ESC self-renewal by regulating a key transcriptional circuit. ESRRB (Estrogen Related Receptor-Beta) and RIF1 (Rap1 Interacting Factor-1 homolog (yeast)) are two main downstream targets in mice. ESRRB belongs to the nuclear hormone receptor superfamily and is essential to prevent differentiation into mesoderm, ectoderm, and neural crest cells, but not trophectoderm differentiation.
RIF1 is an ortholog of a yeast telomeric protein and is upregulated in mouse ES and germ cells. Other genes regulated by Nanog include Tbx3 (T-box 3) and TCL1 (T-Cell Lymphoma breakpoint-1). The function of TCL1 is to repress only a subset of neural crest genes, whereas Tbx3 blocks the mesoderm and ectoderm differentiation. Downregulation of Nanog, SOX2, ESRRB, Tbx3 or TCL1 causes immediate induction of Otx2 [Orthodenticle homolog 2 (Drosophila)], Pitx2 (Paired-like homeodomain Transcription factor-2), Sox18 [SRY (Sex determining region Y)-box 18], and likely other genes. In the epiblast, all three genes are expressed. Oct4, SOX2, and Nanog all suppress the expression of the Caudal-type homeodomain transcription factor Cdx2 (Caudal type homeobox-2). Trophectoderm development is dependent on Cdx2. Oct4, Nanog, and SOX2 bind to roughly 352 genes in undifferentiated human ES cells at the same time, which can be expressed or repressed.
Nanog's ability to maintain ESC identity when overexpressed without the normally required LIF and BMP/GDF growth differentiation factor) signals puts it at the center of the circuitry responsible for self-renewal and pluripotency features. The current understanding of Nanog function and control within these circuits is limited. However, when more genes working upstream and downstream of Nanog are discovered, as well as Nanog's protein partners, a fuller picture of how Nanog works emerges. Nanog's discovery is critical for understanding early embryogenesis and using pluripotent cells for medicinal purposes.
Nanog, a transcription factor with a homeobox domain that regulates embryonic development and cellular reprogramming, has been found to be widely expressed in human malignancies. Nanog is known to have protumorigenic properties, according to functional investigations.
Nanog-mediated oncogenic reprogramming may underpin clinical symptoms of malignant disease, in addition to enhancing self-renewal and long-term proliferative capability of stem-like cancer cells. NANOG enhances tumors' molecular circuitry, and so could be a new therapeutic target or biomarker for cancer diagnosis, prognosis, and therapy outcome.
In conclusion, nanog is a protumorigenic factor that may serve in the clinic as a biomarker for cancer diagnosis, prognosis and predictor of anticancer therapeutic efficacy. Furthermore, NANOG itself may represent a therapeutic target as its elimination is predicted to ablate CSC self-renewal and root out the cause of tumor recurrence and metastasis.
Learn about some of the most frequently asked question about Nanog, pluripotency, and embryonic stem cells.