This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Glutamine is the most abundant and versatile amino acid in the body. In health and disease, immune cells consume glutamine at a rate similar to or higher than glucose.
Glutamine is an amino acid containing five carbon atoms; its molecular weight is 146.15 kDa, and its elemental composition includes carbon (41.09%), hydrogen (6.90%), oxygen (32.84%) and nitrogen (19.17%). In terms of its physiological pH, glutamine is classified as a neutral amino acid, while in terms of nutrition, it is classified as a non-essential amino acid. Glutamine has two amino groups, namely α-amino group and easily hydrolyzed side chain amide group. These characteristics make glutamine can be used as nitrogen transporter and NH3 carrier. Glutamine is also a protein amino acid, that is, the amino acid incorporated into the protein, accounting for 5% to 6% of the combined amino acid.
Synthesis and hydrolysis of glutamine. Glutamine is mainly synthesized by glutamine synthetase (GS) and hydrolyzed by glutaminase (GLS). GS uses glutamate and ammonia (NH3) as sources to catalyze the biosynthesis of glutamine. In this reaction, one ATP is consumed. Glutamate can be provided by many amino acids obtained from exogenous (ie diet) and/or endogenous amino acid catabolism. On the other hand, GLS is responsible for the hydrolysis of glutamine into glutamate and ammonium ions (NH4). Almost all cells in the body express GS and GLS, and their main expression and activity will determine whether the tissue is more likely to produce or consume glutamine.
The amino acid glutamine was first considered to be a biologically important molecule more than 100 years ago, when it was suggested on the basis of indirect evidence that glutamine may occur as a structural component of proteins. It was subsequently shown that free glutamine is abundant in certain plants,’ but relatively little was known about the metabolism of glutamine until the1930s, when it was directly demonstrated that glutamine is a component of proteinS3 and when Hans Krebs first described the capacity for glutamine hydrolysis and glutamine synthesis in mammalian tissues.
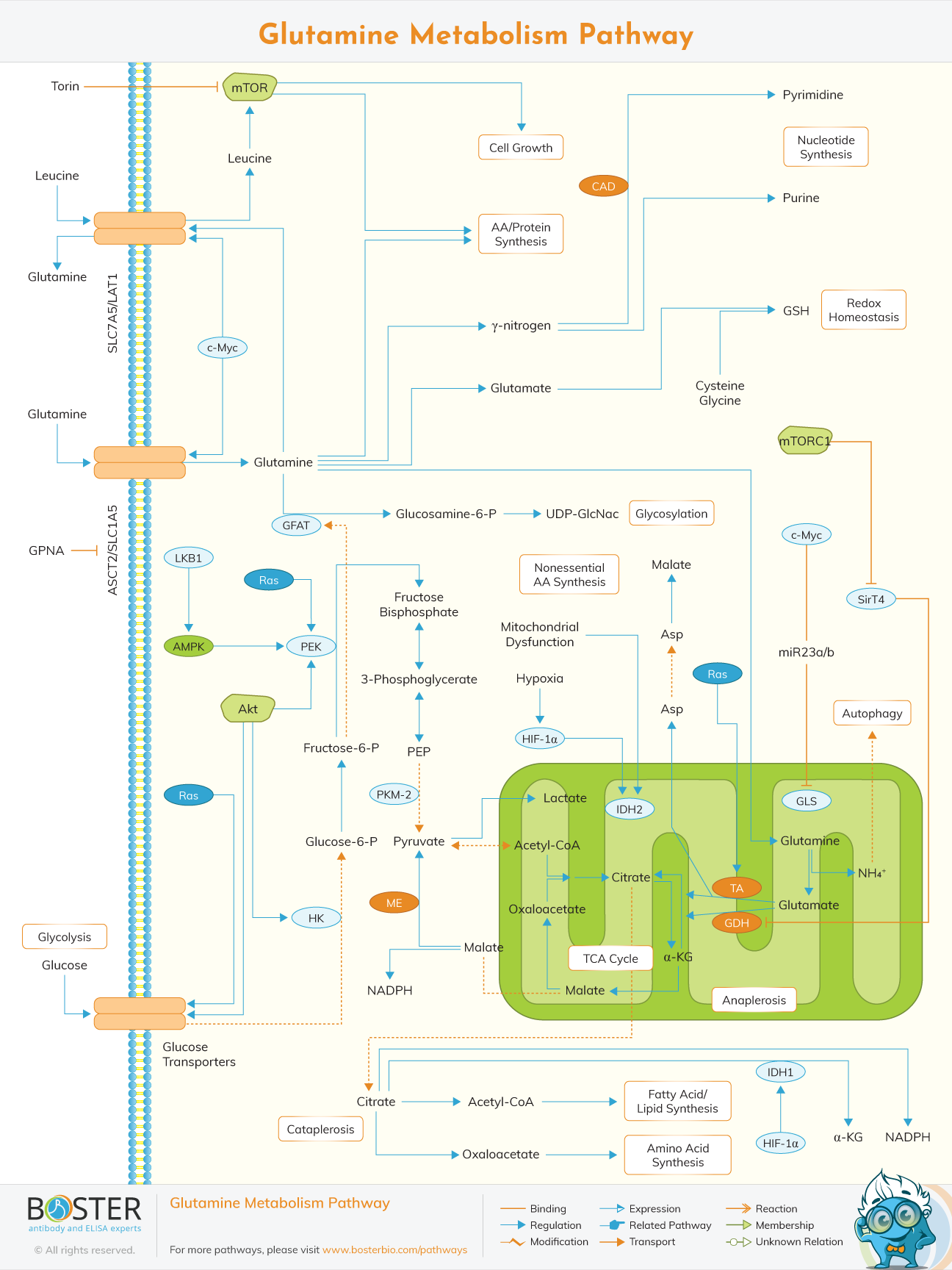
Glutamine is an important metabolic fuel that can help rapidly proliferating cells meet the increasing demand for ATP, biosynthetic precursors, and reducing agents. Glutamine enters the cell via the ASCT2 / SLC1A5 amino acid transporter and is converted to glutamate in the mitochondria through a glutaminase-catalyzed deamination reaction (GLS). Glutamate is converted to intermediate α-ketoglutarate (αKG) of the TCA cycle by glutamate dehydrogenase (GDH) or alanine or aspartate aminotransferase (TA). In addition to αKG, they also produce the corresponding amino acids. ΑKG is a key metabolite that can be used for the production of ATP and the replacement of intermediates in the TCA cycle, a process called replacement. During hypoxia or mitochondrial dysfunction, αKG can be converted to citrate in an IDH2-catalyzed reductive carboxylation reaction.
The newly formed citrate leaves the mitochondria, where it is used to synthesize fatty acids and amino acids and produce the reducing agent NADPH (cataplerosis). In the cytosol, glutamine uses its gamma nitrogen (amide) for the synthesis of nucleotides and hexosamines, and produces glutamate in the process. Cytoplasmic glutamate is essential for maintaining redox homeostasis and protecting cells from oxidative stress through the production of glutathione (GSH).
Many cancer cells show glutamine addiction dependent on oncogenes, and glutamine itself can promote proliferation signals. For example, the entry of glutamine through SLC1A5, combined with its exit through the SLC7A5/LAT1 transporter, allows leucine to enter the cell and trigger mTORC1-mediated cell growth. In addition, Akt, Ras, and AMPK signal molecules activate glycolytic enzymes and induce lactate production (Warburg effect), causing cancer cells to require glutamine metabolism to meet increased energy requirements. The proto-oncogene cMyc up-regulates glutamine breakdown through transcriptional activation of the GLS and SLC1A5 genes. Glutamine-mediated glycosylation of proteins, including growth factor receptors, can direct proteins to the cell surface and induce their activation.
Glutamine uptake is particularly increased in cancer cells with dysregulated tumor suppressor oncogenes (such as cmyc). Glutamine is catabolized by different enzymes, including GLS, CAD, or glutamine fructose 6 phosphate aminotransferase (GFAT). As a source of replenishment, glutamine is converted into ketoglutarate through the breakdown of mitochondrial glutamine. In the irreversible reaction catalyzed by the GLS enzyme, glutamine is first deamidated to glutamate. Then, glutamic acid is deaminated to ketoglutarate by GLUD1/glutamate dehydrogenase (GDH) enzyme or various aminotransferases to produce other non-essential amino acids. Subsequently, ketoglutarate enters the TCA cycle to replenish the mitochondrial citrate pool. GLS is a restriction enzyme that decomposes glutamine, and its regulation is strictly controlled.
There are two isotypes of GLS, which are encoded by two genes in mammals, renal type GLS1 and living type GLS2. GLS1 is the main isotype expressed in cancer cells and has been shown to be upregulated in a variety of cancers, including breast cancer, lung cancer, cervical cancer, and brain cancer. GLS1 is inhibited by its product glutamate. GDH activity is also increased in tumor cells. Leucine is a key amino acid for signal transduction and an allosteric activator of GDH, which can induce the production of α-ketoglutarate and prevent the accumulation of glutamate from inhibiting GLS. As mentioned above, glutamine is imported by the SLC1A5 transporter, while leucine is taken up by the bidirectional antiporter SLC7A5, which exports glutamine to the outside of the cell. Therefore, the combination of glutamine and leucine regulates the breakdown of glutamine.
Glutamine can be synthesized by cells through GLUL / glutamine synthetase (GS), which catalyzes the condensation reaction between glutamate and ammonia in an ATP-dependent manner to produce glutamine.
In mammals, GS is expressed primarily in the liver, brain, and muscle. GS has been found to be a marker for hepatocellular carcinoma (HCC) and its elevated expression may increase the metastatic potential of HCC patients. Furthermore, GS expression is associated with poor survival in glioblastoma patients.
Glutamine metabolism in mammals and the rapamycin complex 1 (mTORC1) pathway are closely related through different mechanisms. The activation of mTORC1 by glutamine and other amino acids is mediated by the Rag GTPase pathway. In addition, glutamine acts as an export solute for leucine input, supporting glutamine to activate mTORC1 through glutamine decomposition. In addition, glutamine and leucine cooperate to produce ketoglutarate through the decomposition of glutamine, which ultimately activates mTORC1. In fact, short-term glutamine breakdown induces lysosomal translocation and mTORC1 activation through Rag GTPase, then inhibits autophagy and promotes cell growth.
Consistent with this positive correlation between glutamine breakdown and mTORC1, FOXO-mediated GS expression inhibits mTOR signaling by blocking its lysosomal translocation. This mechanism is important for maintaining autophagy during periods of nutritional deficiency. Therefore, mTORC1 detects the availability of glutamine in two directions: when glutamine is available, mTORC1 is activated by the production of α-ketoglutarate; but when glutamine production is activated, mTORC1 is inactivated.
In view of the dependence of cancer cells on glutamine metabolism, targeted therapies for glutamine metabolism have been developed, from glutamine uptake to glutamine catalytic enzyme. Due to the dysregulation of GLS in a variety of cancers, GLS inhibition has attracted attention.
In addition to GLS inhibitors, strategies that target the conversion of glutamate to α-ketoglutarate, such as GDH inhibitors and transaminase inhibitors, have also been evaluated in preclinical models of breast cancer and neuroblastoma..