This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
The process of glucose conversion to lactate even under normal oxygen levels in tumors cells.
Otto Warburg and colleagues discovered in the 1920s that tumors absorbed huge quantities of glucose in comparison to surrounding tissue. Furthermore, even in the presence of oxygen, glucose was digested to generate lactate, giving rise to the phrase aerobic glycolysis. However, it was also shown that tumor viability might be maintained solely by breathing.
As a result, it was determined that in order to destroy tumor cells by depriving them of energy, both glucose and oxygen must be removed . Following that, in 1929, an English scientist named Herbert Crabtree expanded on Warburg's findings by studying the heterogeneity of glycolysis in tumor types. He validated Warburg's results, but also showed that the intensity of respiration in tumors varied exhibiting a substantial amount of respiration.
Warbug later came up a proposal that dysfunctional mitochondria is the root of aerobic glycolysis, which he hypothesized as the primary cause of cancer. In 1970, this phenomenon was termed as The Warbug effect by Efraim Racker.
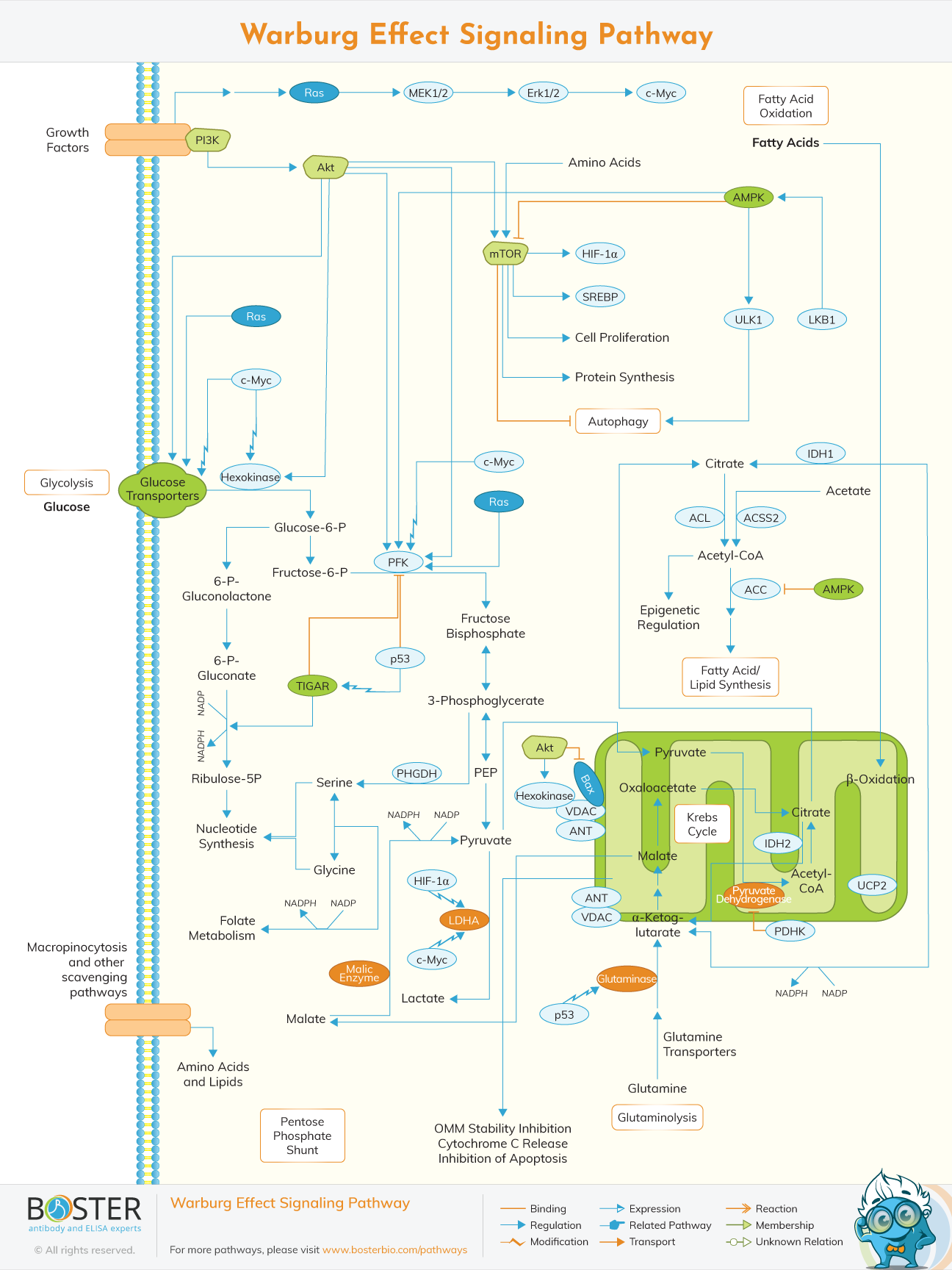
The Warburg effect is a metabolic phenotype commonly seen in tumors. Even in the presence of sufficient oxygen, cancer cells produce about 60% of their ATP through glycolysis instead of oxidative phosphorylation. The Warburg effect allows dividing cells to use intermediate glucose metabolites to double their biomass and suppress apoptosis. This phenomenon was first discovered by Otto Warburg in the 1920s and was called aerobic glycolysis.
Unlike normal differentiated cells, which mainly rely on mitochondrial oxidative phosphorylation to generate energy for cellular processes, cancer cells reconnect their metabolism through aerobic glycolysis to promote growth, survival, proliferation and long-term maintenance. The common feature of this metabolic change is increased glucose uptake and fermentation of glucose to lactic acid. This phenomenon can be observed even in the presence of fully functional mitochondria, and is collectively referred to as the Warburg effect.
The metabolic circuit allows the production of NADPH and the flow of acetyl-CoA to the cytoplasm for the synthesis of lipids. Activation of growth factor receptors leads to tyrosine kinase signaling and PI3K activation. Through AKT, PI3K activation stimulates glucose uptake and flow through the early part of glycolysis. cMyc boosts glutamine metabolism, which also supports NADPH production. LKB1 / AMPK and p53 signaling reduce metabolic flux through glycolysis in response to cellular stress. The decrease in glycolytic flux in response to LKB1/AMPK or p53 may be an adaptive response that stops proliferation and metabolism during periods of low energy availability or oxidative stress.
Growth factor signaling also regulates the activity of pyruvate kinase M2 (PKM2) through tyrosine kinases and regulates carbon flux through subsequent glycolysis steps. This regulation of pyruvate kinase can promote the redirection of glucose metabolites to the pentose phosphate shunt, as well as nucleotide and amino acid biosynthesis pathways. The conversion of glucose and glutamine to lactic acid involves lactate dehydrogenase A.
Mitochondrial uptake of glutamine is essential for lipid synthesis because it provides carbon in the form of mitochondrial oxaloacetate to maintain citric acid production during the first step of the TCA cycle. Mitochondria are a key component of the biosynthetic process. The substrates in the TCA cycle (such as malic acid, oxaloacetic acid, citric acid, α-ketoglutarate, and succinic acid) are used for nucleotide, amino acid, and lipid synthesis. Biosynthesis.
The Warburg effect confers direct signaling functions on tumor cells. Glucose can be transported into the plasma by glucose transporter and is phosphorylated into glucose 6 phosphate (G6P) by hexokinase, which is regulated by cMyc and Akt. G6P is converted to fructose 6 phosphate (F6P), which is further phosphorylated to fructose 1,6 diphosphate (FBP) under phosphofructokinase (PFK) for glycolysis. The cleavage products of FBP are glyceraldehyde phosphate (G3P) and dihydroxyacetone phosphate (DHAP). It was originally reported that phosphoenolpyruvate (PEP) partially inhibited FBP activity at high concentrations.
G6P can also be used to synthesize nucleotides through the dependent form of NADP + / NADPH and the enzyme glucose 6 phosphate dehydrogenase (G6PDH) to produce 6P gluconolactone, 6P gluconic acid and ribose 5 phosphate.
Macropinocytosis is a process that allows a large amount of extracellular material that can be transported to the lysosome to be absorbed. It is a way for cells to break down extracellular material and provide nutrients for cell metabolism. These nutrients can produce ATP or NADPH, or directly contribute to biomass. There is also evidence that some cancer cells can scavenge extracellular protein, amino acids, and lipids.