This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
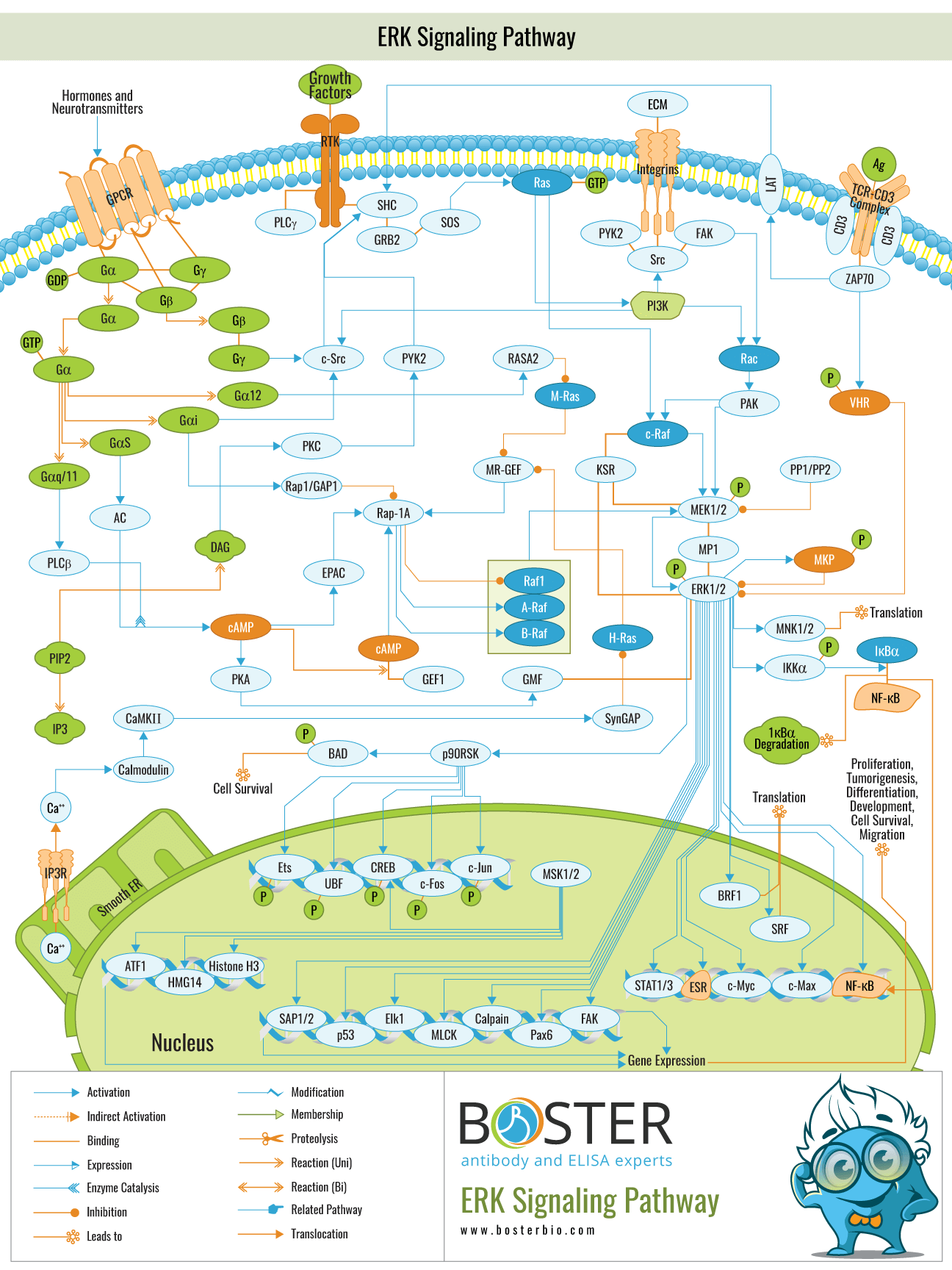
The extracellular signal-regulated kinase (ERK) pathway is a key component of the mitogen-activated protein kinase (MAPK) signaling pathway. The ERK cascade is activated by a variety of extracellular agents, including growth factors, hormones, and cellular stresses, to induce a variety of cellular processes, primarily proliferation and differentiation, but also stress response and others under certain conditions. The major components of MAPK/ERK kinase kinase (MEKK) are the Raf family members.
Raf-1, A-Raf, and B-Raf, which phosphorylate two serine residues on MAPK/ERK kinase (MEK) components MEK1/2. ERK1/2 stimulate downstream effectors, the majority of which are transcription factors. The cyclic AMP response element-binding protein is a significant transcription factor (CREB).
The ERK1/2 cascade is primarily activated by membrane receptors, including receptor Tyr kinases (RTKs), G protein–coupled receptors (GPCRs), and ion channels. These receptors transmit the signal by recruiting adaptor proteins (e.g., Grb2) and exchange factors (e.g., SOS), which activate Ras at plasma membranes or other organelle membranes. The activated, GTP-bound Ras then transmits the signal by activating the Raf-1, B-Raf, and A-Raf (Rafs) protein kinases within the MAP3K level of this cascade. This activation is accomplished by attracting Rafs to membranes, where they are phosphorylated and activated. MOS is another MAP3K in the ERK cascade, but it is primarily expressed in the reproductive system and is regulated differently. The signal is then transmitted down the cascade through the MAPKK components MEK1/2 from the MAP3K level. They are activated via serine phosphorylation of the Ser-Xaa-Ala-Xaa-Ser/Thr motif in their activation loop, which is characteristic of MAPKKs. MEK1/2 then activate their only known substrates, native ERK1/2, which act as their sole downstream targets, implying that MEK1/2 act as the cascade's specificity-determining components. MEK1/2 are the only dual-specificity protein kinases capable of phosphorylating ERK1/2's regulatory Thr and Tyr residues. To date, approximately 200 distinct ERK1/2 substrates have been identified. They are either cytosolic substrates (e.g., PLA2 RSK) or nuclear substrates (e.g., transcription factors such as Elk1, c-Fos, and c-Jun).
At the MAP3K level of the cascade, Raf-1 is the best studied of the growth factor signaling molecules. It is a 70–75 kDa serine/threonine kinase that contains a kinase domain in the COOH-terminal half and a regulatory domain in the remainder of the molecule. Raf-1 is transiently activated within 2-3 mm in response to various mitogens. The mechanism by which Raf-1 is activated has been extensively studied over the last few years. Raf-1 is now known to be located downstream of HAS, which appears to interact directly with the Raf-1 NH2-terminal region upon stimulation.
MEKs are an evolutionary conserved family of serine/threonine protein kinases that currently consists of three highly homologous (85%) mammalian isoforms. MEK-1 activation appears to be mediated solely by serine residue phosphorylation and appears to be similar whether phosphorylated by MEKK, Raf-1, or autophosphorylation, as determined by phosphopeptide maps. MAPKKs (MEKs) are highly specific for the ERK-1 and ERK-2 downstream components. Additionally, MAPKK was unable to recognize denatured MAPK or peptides containing ERK phosphorylation sites, indicating that the enzyme requires the native form of MAPK.
ERK-1 and ERK-2 are frequently considered functionally redundant due to their high degree of similarity. MEK phosphorylation is the primary upstream mechanism that results in the phosphorylation of ERK-1 and -2 on both of these regulatory residues. Due to the ERKs' relatively broad substrate recognition, they can phosphorylate a large number of proteins following activation. Often regulatory in nature, these proteins are found in both the cytoplasm and the nucleus.
Regulating Erk signaling is critical for cells to function normally. The cell can regulate the Erk signaling pathway in a variety of ways. These include regulation via feedback loops, upstream and downstream scaffolds, phosphatase, and inhibitors of the Erk signaling pathway.
The ERK pathway contains several points of negative feedback . MEK is one such target that can be inhibited by ERK-mediated phosphorylation of MEK1/2 at Thr292 and Thr212. Phosphorylation prevents PAK1 from further enhancing MEK activity, thereby inhibiting ERK activation. A second possible feedback loop mechanism is the phosphorylation of Raf at multiple sites by ERK. Hyperphosphorylation of these sites prevents Raf from interacting with the Ras GTPase and promotes dephosphorylation of Raf by the phosphatase PP2A. ERK can also act as a negative feedback mechanism by activating or upregulating the transcription of phosphatases. These various ERK-activated negative feedback loops are one way to limit the duration of the signal and restore the pathway to its basal state.
Scaffold proteins act as a bridge between two or more components of a signaling pathway, bringing them closer together and facilitating their functional interaction. Additionally, scaffolds can direct these multi-enzyme signaling modules to specific cellular locations, enhancing phosphorylation of a subset of downstream substrates. Additionally, scaffolds affect the signal's duration, prevent crosstalk between similar pathways, and can link the signaling module's activation to a specific upstream activating signal.
The MAPK pathways' activity is determined by the competition between upstream kinases and inhibitory phosphatases. The MAPK pathways are terminated when their components are dephosphorylated by a variety of phosphatases. Numerous serine/threonine phosphatases, including PP2A and PP2Ca, and tyrosine phosphatases, including PTP-SL and HePTP, have been demonstrated to inhibit MAPKs.