This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
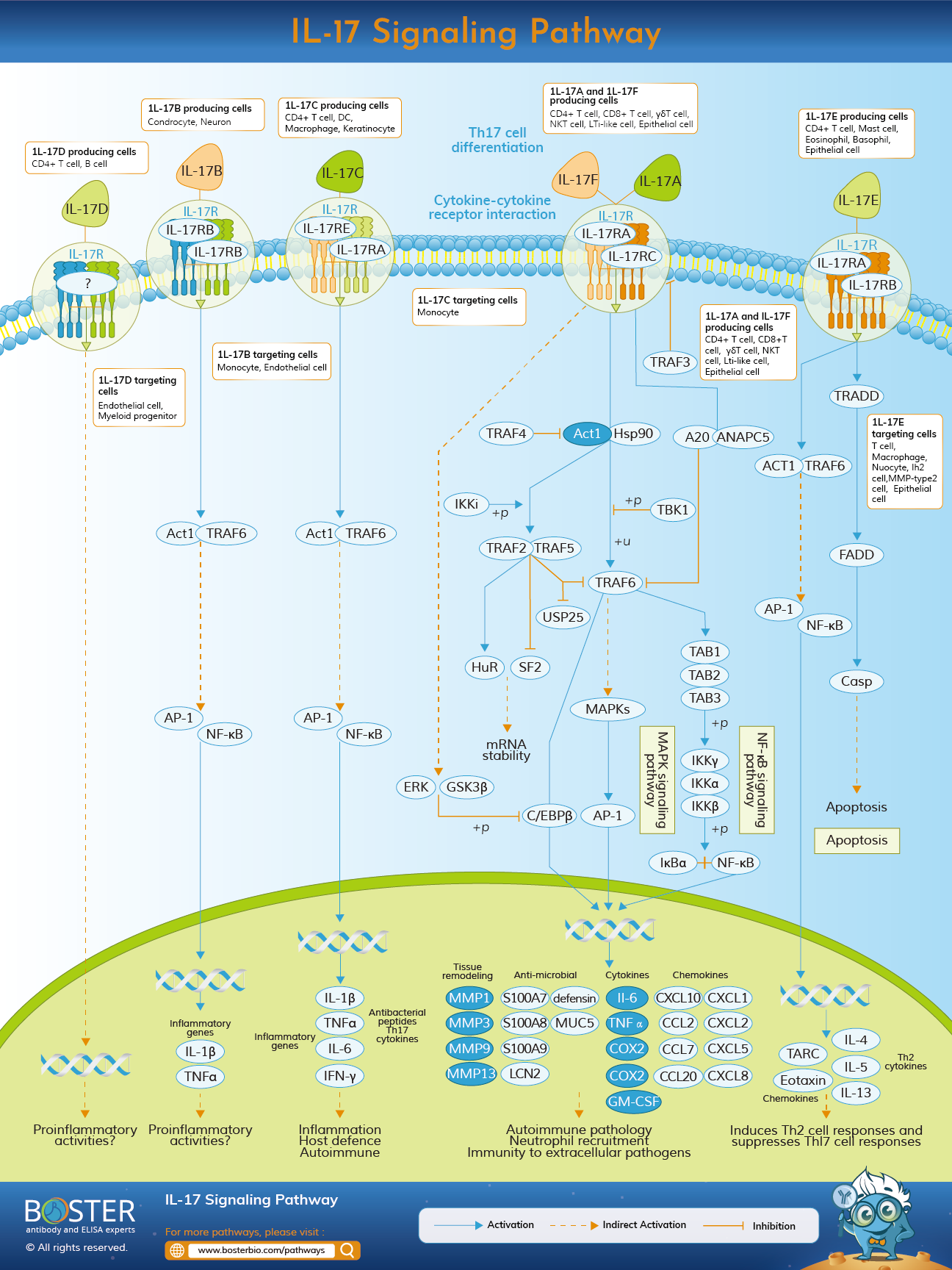
The IL-17 family of cytokines consists of six members: IL-17/IL-17A, IL-17B, IL-17C, IL-17D, IL-17E/IL-25, and IL-17F. These cytokines are produced by a variety of cell types and are primarily involved in the promotion of pro-inflammatory immune responses. Cytokines in the IL-17 family are secreted in dimer form and interact with members of the IL-17 receptor family
The IL-17 receptor family consists of five members: IL-17 R/IL-17RA, IL-17RB, IL-17RC, IL-17RD/SEF, and IL-17RE. Each of these receptor subunits is a type I transmembrane protein with two extracellular fibronectin-like domains and a cytoplasmic SEF/IL-17 R/TIR (SEFIR) domain.
The IL-17 signaling pathway plays a role in the host's defense against extracellular bacterial and fungal infections, as well as the onset of several autoimmune diseases.
Unlike the other members of the IL-17 family, IL-17RA contains a cytoplasmic Toll/IL-1 R-like loop (TILL) as well as a C/EBP beta activation domain (CBAD). The receptors of the IL-17 family oligomerize to form functional receptor complexes. Although the stoichiometry of these complexes is unknown, it is believed that the receptor for IL-17A, IL-17F, and IL-17A/F is a heterotrimeric complex composed of two IL-17RA subunits and one IL-17RC subunit. Additionally, the IL-17RA receptor subunit interacts with the IL-17RE and IL-17B R/IL-17RB to form functional receptor complexes for IL-17C and IL-17E/IL-25. IL-17 is required for proper host immunology. Similar to IL-17C, IL-17A is a hallmark of this family in that it protects the host against extracellular pathogens while also promoting inflammatory pathology in autoimmune disease. While IL-17F plays a role in mucosal host defense, IL-17E boosts the Th2 immune response. Antibacterial cytokines and chemokines are activated by the IL-17 family via the MAPK, NF-B, and C/EBPs pathways.
TRAF6 was one of the first mediators identified in the 1L-17RA signaling pathway. Schwandner R et al. discovered that TRAF6 is required for II-17A to activate NF-Kappa B and JNK, and that IL-17A cannot induce IL-6 production in TRAF6-deficient mouse embryonic fibroblasts (MEFs). However, there is no obvious TRAF6 binding site in IL-17RA, implying that another mediator acts as a bridge between IL-17RA and TRAF6.
IL-17 activates Erk by phosphorylating Thr188 of C / EBP beta via the SEFIR-TILL domain of IL-17RA. Phosphorylation of Thr188 is also required for GSK 3 beta to phosphorylate Thr179 of C / EBP beta. These two sequential phosphorylation events result in C / EBP beta inactivation. Additionally, TRAF3 inhibits IL-17 signal transduction and the induction of downstream inflammatory factors by competitively binding to IL-17R.
In 2003, Novatchkova M. identified a conserved domain in the intracellular segment of the IL-17R family using bioinformatics analysis and named it SEFIR. Simultaneously, an intracellular protein Act1 was discovered to contain the SEFIR domain. Act1 was originally named for its role as a mediator of NF-kappa B activation. Recent studies, however, have confirmed that Act1 acts as a linker between IL-17RA and TRAF6. Act1 is regarded as the pathway's master mediator. It is capable of interacting with IL-17R via the SEFIR domain. And Act1 contains the TRAF6 binding site. Through Act 1 gene silencing or RNA interference, IL-17R is required to activate NF-kappa B. Additional studies revealed that Act1, as a U-box E3 ligase, modifies TRAF6 at K124 via K63 polyubiquitination, thereby mediating IL-17R downstream signaling and inflammatory gene expression.