This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
The process by which epithelial cells differentiate into motile mesenchymal cells is critical for development, wound healing, and stem cell behavior, and contributes pathologically to fibrosis and cancer progression.
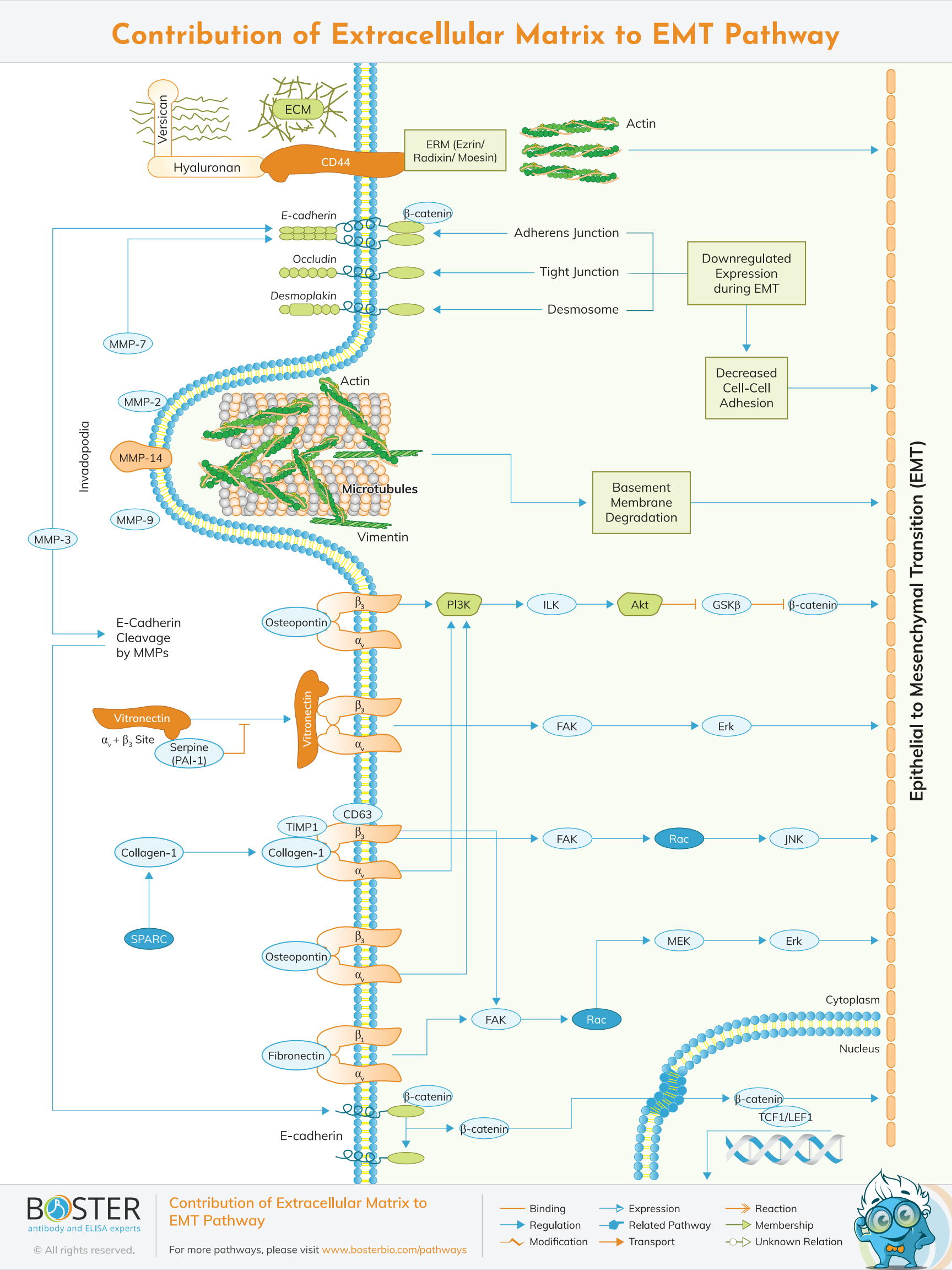
Extracellular matrix is the non-cellular component present within the tissue and organs and provides not only essential physical scaffolding for the cellular constituents, but also initiates crucial biochemical and biomechanical cues that are required for tissue morphogenesis,differentiation and homeostatis. EMT on the other hand is the integral in development,wound healing and stem cell behaviour and contributes pathologically to fibrosis and cancer progression.
Epithelial to mesenchymal transition (EMT) cells acquire enhanced migratory capabilities. To migrate from one location to another, a cell must first invade the surrounding tissue via localized regions of disassembled basement membrane. Invadopodia are specialized regions of a cell that protrude through the basement membrane and initiate invasion in response to pathological conditions. Osteoclasts contain related structures called podosomes during the physiological process of bone remodeling. Invadopodia are Actin-rich membrane protrusions that accumulate MMPs required for extracellular matrix (ECM) remodeling and are thought to contribute to the in vivo early stages of metastasis.
It was first outlined as a feature of embryogenesis in 1980s by Betty Hay. Epithelial and mesenchymal cells differ phenotypically as well as function but they both share inherent plasticity. Epithelial are closely connected to each other by gap junctions, adherens and tight junctions, polarization of the action cytoskeleton have an apical-basal polarity and bound by a basal lamina. Mesenchymal cells in contrary lack this polarization. They have a spindle-shaped morphology and interact with each other through focal points only.
One of the defining characteristics of EMT is the loss of epithelial integrity, which occurs as a result of the degradation of adherens junctions that maintain epithelial cell-cell contact. Proteolytic digestion by matrix metalloproteinases (MMPs), the expression of which is regulated by EMT-associated signaling pathways (e.g., TGF-), is a primary driver of this degradation. The degradation of adherens junctions is further facilitated by pro-EMT transcription factors such as SNAIL repressing genes encoding epithelial-specific proteins (e.g., E-Cadherin, Occludin, Desmoplakin). During EMT, certain components of epithelial adherens junctions are replaced with proteins (e.g., N-Cadherin) that increase junctional flexibility, allowing for cell detachment and increased motility. EMT also involves remodeling of the action cytoskeleton, which is facilitated in part by altered expression of Ezrin/Radixin/Moesin (ERM) proteins such as Moesin in invasive breast cancer cells. Additionally, ERM proteins interact with CD44, a cell surface glycoprotein that is abundantly expressed in some cancer stem cells and is strongly associated with cell motility and metastasis, in part due to its function as a receptor for extracellular proteoglycans such as hyaluronan and versican.
E-cadherin is in its high levels in the epithelial cells while N-cadherin,fibronectin and vimentin are in their high levels in mesenchymal cells. E-cadherin loss is considered to be a fundamental event in EMT. Transcriptional factors that repress E-cadherin directly or indirectly are considered as EMT-TF. SNA1,2,ZEB1,2,TCF3 and KLF-8 can bind to the E-cadherin promoter and repress its transcription, whereas Twist,TCF-4 repress E-canderin indirectly.
ZEB and SNAIL factors bind to the E-box consensus sequence on the promoter region,while KLF8 binds to the promoter through GT boxes. EMT-TF repress transcriptionally other junctional proteins like claudins and desmosomes,thus enhancing EMT.Many EMT-TF are involved in promoting metastatic events due to the fact that EMT in cancer progression recaptures EMT in developmental programs. Some signalling pathways like the TGF beta,FGF,EGF,HGF, beta-catenin and notch and hypoxia may induce EMT.
Twist has been shown to induce the formation of actin-rich membrane protrusions called invadopodia, which recruit the proteases MMP-7, MMP-9, and MMP-14 to the leading edge, where they degrade the ECM and basement membranes, thereby facilitating tumor invasion and metastasis. Twist, Slug, Snail, and Zeb have been shown to upregulate the expression of several epithelial extracellular matrix (ECM) proteins during EMT, including collagen 1, vitronectin, and fibronectin. Several integrin complexes are also upregulated during EMT, including integrin 51, which binds fibronectin, and integrins 11 and 21, which interact with collagen I and have been shown to mediate E-cadherin complex disruption. ECM-associated proteins such as SPARC, a glycoprotein that promotes the interaction of collagen and 21, have been shown to modulate cellular interactions with the ECM.
SPARC induces EMT by modulating the expression of SLUG and has been linked to increased aggressiveness in melanoma. SERPINE1 (PAI-1), another component of the epithelial ECM, inhibits vitronectin's binding to integrin v3. Serpin levels have been associated with aggressiveness in a variety of cancer types; this has been attributed to Serpin-mediated disruption of cell-ECM interfaces. TIMP1, in conjunction with CD63, has been shown to mediate an EMT-like transformation of normal cells by increasing the activity of 1 integrin signaling.