This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Nuclear factor-κB (NF-κB) function as dimeric transcription factors that regulate the expression of genes influencing a broad range of biological processes including innate and adaptive immunity, inflammation, stress responses, B-cell development, and lymphoid organogenesis.
Nuclear Factor kappa-light-chain-enhancer of activated B cells (NF-kB) is a short name for Nuclear Factor kappa-light-chain-enhancer of activated B cells. It is a small family of inducible transcription factors that play a significant role in practically all mammalian cells, rather than a single protein. It regulates DNA transcription, cytokine synthesis, cell survival, and other key cell activities, and is particularly important in regulating the immunological response to infection.
NF-kB dimers are the most common type of NF-kB molecules. P50-P65 dimer (NF-kB1/RelA) is a common NF-kB structure. The development of a dimer is required for DNA binding because two NF-B monomers bind to DNA as a dimer.
The dimer's N-terminal domains are in charge of particular DNA interaction. The C-terminal regions of proteins are usually highly conserved; they are involved in dimerization and nonspecific DNA phosphate interaction. The entire NF-kB molecule functions as a transcription factor and acts like a pliers vise on the DNA chain.
NF-B family members are also known as NF-kB/Rel Proteins because they share structural homology with the retroviral oncoprotein v-Rel. In mammals, there are five proteins in the NF-kB transcription factor family: p65/RelA, RelB, c-Rel, p105/p50 (NF-kB1), and p100/p52 (NF-kB2). These five proteins produce unique transcriptionally active homo/heterodimeric complexes when they interact with one another . They all have a 300-aa Rel homology domain (RHD) that is conserved. Dimerization, DNA binding, interaction with IkBs, and nuclear translocation are all functions of the RHD domain. Although members of the NF-kB protein family can create up to 15 distinct dimers, many of these have yet to be discovered. The p50/p65 heterodimer, which has been found in almost all cell types, is the most common form of NF-kB dimer.
The NF-kB family of proteins can be separated into two groups because only P65/Rel, RelB, and c-Rel have carboxy-terminal transactivation domains (TAD). The precursor molecules p105 and p100 are processed to produce p50 and p52, respectively. As a result, not all Rel dimer combinations are transcriptionally active.
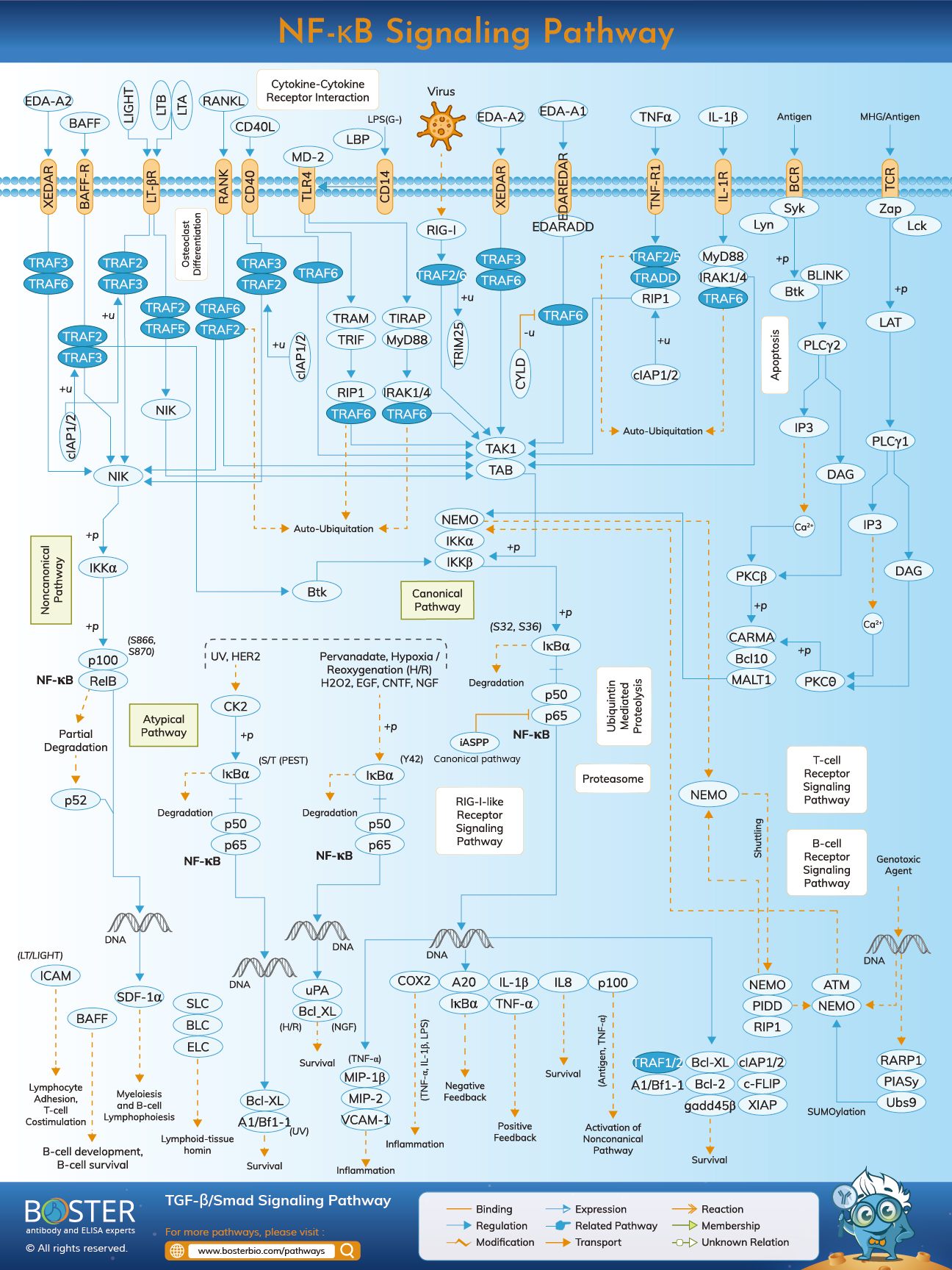
As nuclear transcription factors, NF-kB dimmers must travel to the nucleus and interact with DNA in order to function. NF-KB was inactive and retained in the cytoplasm of the majority of normal cell types during the resting state. They attach to a particular inhibitor termed IK-B protein, which may interact with NF-Rel kB's homology domain (RHD) and impair its nuclear localization sequence (NLS) activity. IkBa, IkBb, and IkBg are inhibitor proteins that include 6–7 ankyrin repeats that mediate binding to the RHD. These repeats are also present in the C-terminal portions of the NF-kB2/p100 and NF-kB1/p105 precursors, which act as IkBs and maintain their cytoplasmic partners, the Rel proteins. To activate the NF-kB molecular pathway, cells must first dissociate the NF-kB protein from its inhibitors. Two primary signaling pathways result in the dissociation of the IK-B protein inhibitor from NF-kB dimers and the translocation of NF-kB dimers from the cytoplasm to the nucleus.
To activate the NF-kB molecular pathway, cells must first dissociate the NF-kB protein from its inhibitors. Two primary signaling pathways result in the dissociation of the IK-B protein inhibitor from NF-kB dimers and the translocation of NF-kB dimers from the cytoplasm to the nucleus.
Cascade signaling initiated by pro-inflammatory cytokines and pathogen-associated molecular patterns (PAMPs) on the cell surface, such as the tumor necrosis factor receptor (TNFR), toll-like receptor (TLR), and T/B cell receptor. When these receptors attach to their ligand molecules and transmit the signal across the cell membrane, the IkB kinase (IKK) complex is activated. This complex is most frequently found as a heterodimer composed of IKK and IKK catalytic subunits and an IKK regulatory subunit. The IKK unit is also known as NEMO, which stands for NF-kB essential modulator.
The active IKK complex catalyzes the phosphorylation of IkBs (at Ser32 and Ser36 of IkBa), polyubiquitination (at Lys21 and Lys22 of IkBa), and subsequent destruction by the 26S proteasome. The released NF-kB dimers (most frequently the p50–RelA dimer) translocate to the nucleus, bind to DNA, and stimulate transcription of downstream genes.
Another mechanism that activates NF-kB is referred to as the non-canonical or alternative pathway. This pathway is not dependent on IKK and IKK, but on IKK dimer. Transfer of signals into the cytoplasm via the LT- or BAFF receptor. Phosphonate an NIK protein, and then the IKK homodimmers. In this pathway, IKK homodimers bind to NF-kB2/p100, a protein that is phosphorylated at two C-terminal locations. Phosphorylation of these sites is required for the conversion of p100 to p52, which is likewise polyubiquitinated and degraded by the proteasome. However, unlike with IkBs, phosphorylation-dependent ubiquitination of p100 leads in the destruction of only its inhibitory C-terminal half portions.
Once the C-terminal half of NF-kB (the p52 polypeptide containing the RHD) is degraded, the N-terminal half is released. Due to the fact that the RHD of p100 is frequently linked with RelB, activation of this 'alternative' pathway results in nuclear translocation of p52–RelB dimers. Finally, the dimer binds to DNA and activates downstream gene transcription.
Ubiquitin is a 76-amino-acid protein that is highly conserved and is expressed ubiquitously in all eukaryotes, from yeast to humans. Through an isopeptide bond, the carboxylic acid in ubiquitin's C-terminal glycine can be covalently connected to the epsilon amine of lysine on another protein. This is referred to as "ubiquitination." Additionally, the C-terminal tail of ubiquitin can be directly connected to another ubiquitin's N-terminal methionine to produce a linear polyubiquitin chain. Ubiquitination is required for the proper regulation of NF-kB pathways. NF-kB is sequestered in the cytoplasm of unstimulated cells by binding to inhibitory proteins of the kB family (IkB). IkB is phosphorylated upon activation by the IkB kinase (IKK) complex and is then ubiquitinated and destroyed by the 26S proteasome, allowing NF-kB to translocate to the nucleus, where it regulates the expression of a large number of genes. As the IKK complex's regulatory member, NEMO has been postulated to be the critical factor in transducing the ubiquitination signal to IKK activation. Numerous DUBs act as critical negative regulators of IKK, allowing for precise control of NF-kB activation. A20 is one of the most extensively researched DUBs.
It has been postulated that A20 inhibits NFkB hyperactivation by deubiquitinating K63-linked polyubiquitin bound to RIP1 via the N-terminal OTU domain and encouraging K48-linked polyubiquitination of RIP1 for proteasomal destruction.