This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

3 Citations 17 Q&As



1 Citations
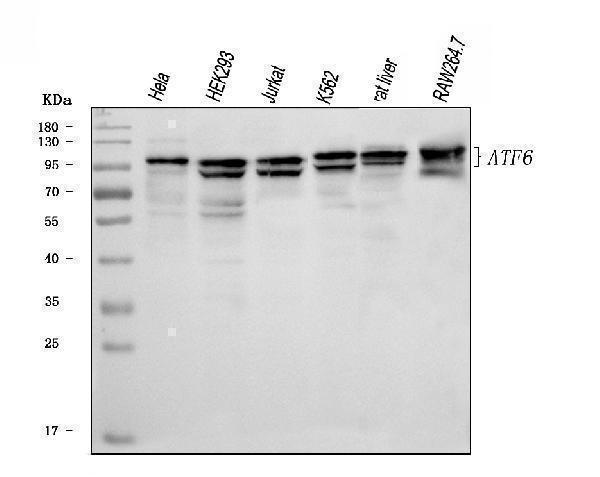
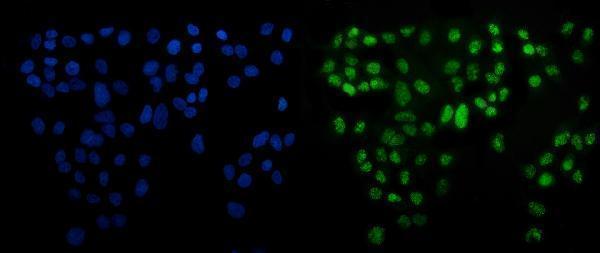
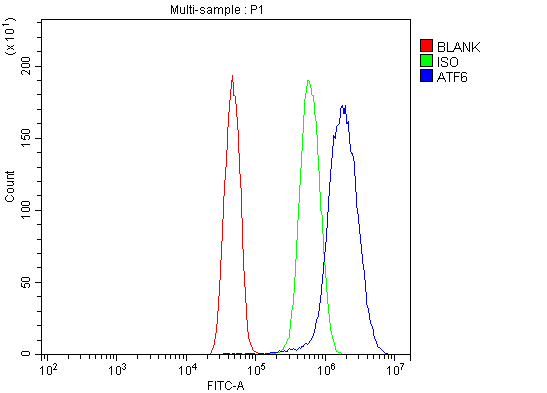
Facts about Cyclic AMP-dependent transcription factor ATF-6 alpha.

Binds DNA on the 5'-CCAC[GA]-3'half of the ER stress response element (ERSE) (5'-CCAAT-N(9)-CCAC[GA]-3') and of ERSE II (5'- ATTGG-N-CCACG-3'). Binding to ERSE requires binding of NF-Y to ERSE.
| Human | |
|---|---|
| Gene Name: | ATF6 |
| Uniprot: | P18850 |
| Entrez: | 22926 |

| Belongs to: |
|---|
| bZIP family |

ACHM7; Activating transcription factor 6 alpha; activating transcription factor 6; atf6 a; ATF6 alpha; ATF6; ATF6A; ATF6-alpha; cAMP-dependent transcription factor ATF-6 alpha; cyclic AMP-dependent transcription factor ATF-6 alpha
Mass (kDA):
74.585 kDA

| Human | |
|---|---|
| Location: | 1q23.3 |
| Sequence: | 1; NC_000001.11 (161766320..161964070) |
Ubiquitous.
Endoplasmic reticulum membrane; Single-pass type II membrane protein.; [Processed cyclic AMP-dependent transcription factor ATF-6 alpha]: Nucleus. Under ER stress the cleaved N-terminal cytoplasmic domain translocates into the nucleus. THBS4 promotes its nuclear shuttling.





PMID: 9271374 by Zhu C., et al. Interaction of ATF6 and serum response factor.
PMID: 9837962 by Yoshida H., et al. Identification of the cis-acting endoplasmic reticulum stress response element responsible for transcriptional induction of mammalian glucose-regulated proteins; involvement of basic-leucine zipper transcription factors.
*More publications can be found for each product on its corresponding product page