This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

Facts about Potassium voltage-gated channel subfamily KQT member 5.

May contribute, together with other potassium channels, to the molecular diversity of a heterogeneous population of M- channels, varying in kinetic and pharmacological properties, which underlie this physiologically significant current. Insensitive to tetraethylammonium, but inhibited by barium, linopirdine and XE991.
| Human | |
|---|---|
| Gene Name: | KCNQ5 |
| Uniprot: | Q9NR82 |
| Entrez: | 56479 |

| Belongs to: |
|---|
| potassium channel family |

KQT-like 5; Kv7.5; potassium channel protein; Potassium channel subunit alpha KvLQT5; potassium voltage-gated channel subfamily KQT member 5; potassium voltage-gated channel, KQT-like subfamily, member 5; Voltage-gated potassium channel subunit Kv7.5
Mass (kDA):
102.179 kDA

| Human | |
|---|---|
| Location: | 6q13 |
| Sequence: | 6; NC_000006.12 (72621843..73198853) |
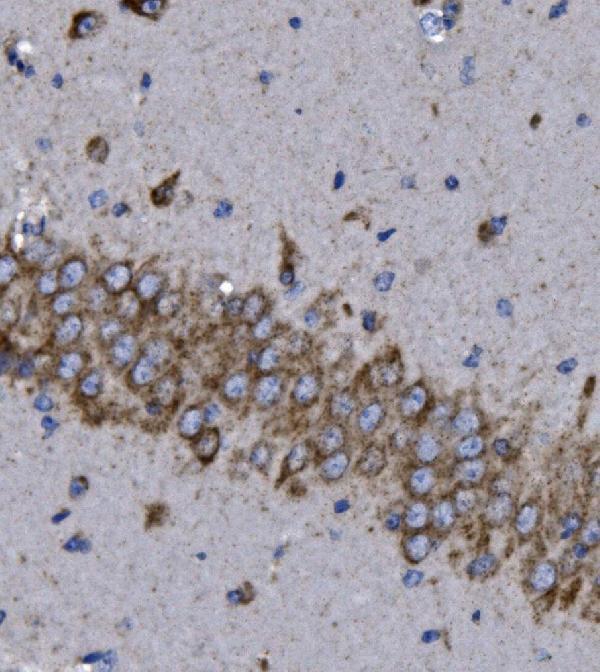
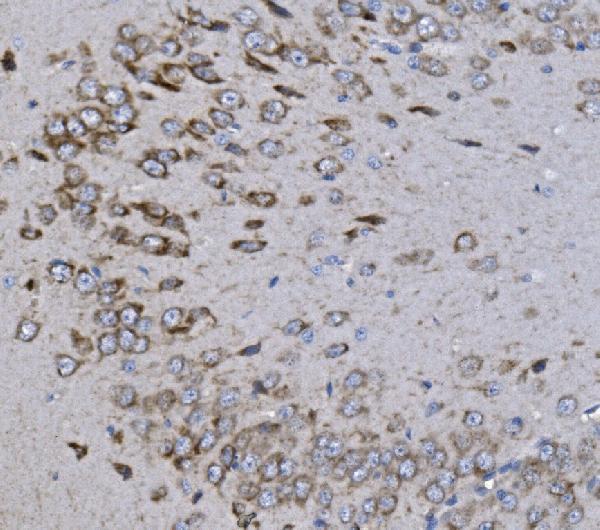
Strongly expressed in brain and skeletal muscle. In brain, expressed in cerebral cortex, occipital pole, frontal lobe and temporal lobe. Lower levels in hippocampus and putamen. Low to undetectable levels in medulla, cerebellum and thalamus.
Cell membrane; Multi-pass membrane protein.



PMID: 10787416 by Lerche C., et al. Molecular cloning and functional expression of KCNQ5, a potassium channel subunit that may contribute to neuronal M-current diversity.
PMID: 10816588 by Schroeder B.C., et al. KCNQ5, a novel potassium channel broadly expressed in brain, mediates M-type currents.