This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

1 Citations 1 Q&As

Facts about Homeobox protein Nkx-2.5.

Binds to the core DNA motif of NPPA promoter (PubMed:22849347, PubMed:26926761). It is transcriptionally controlled by PBX1 and acts as a transcriptional repressor of CDKN2B (By similarity).
| Human | |
|---|---|
| Gene Name: | NKX2-5 |
| Uniprot: | P52952 |
| Entrez: | 1482 |

| Belongs to: |
|---|
| NK-2 homeobox family |

cardiac-specific homeo box; cardiac-specific homeobox 1; Cardiac-specific homeobox; CHNG5; CSX1; CSXFLJ97195; FLJ52202; FLJ99536; Homeobox protein CSX; Homeobox protein NK-2 homolog E; homeobox protein Nkx-2.5; NK2 transcription factor related, locus 5 (Drosophila); NKX2.5; NKX2.5FLJ97166; NKX2-5; NKX2E; NKX2EFLJ97197; NKX4-1; tinman paralog
Mass (kDA):
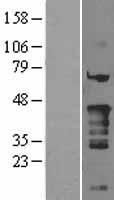
34.918 kDA

| Human | |
|---|---|
| Location: | 5q35.1 |
| Sequence: | 5; NC_000005.10 (173232109..173235321, complement) |
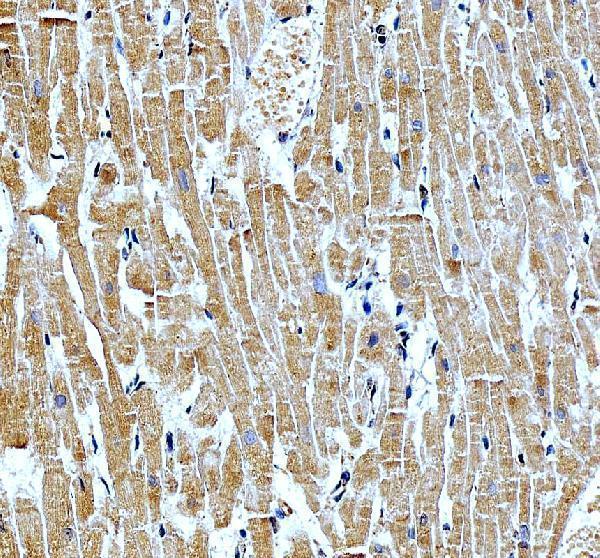
Expressed only in the heart.
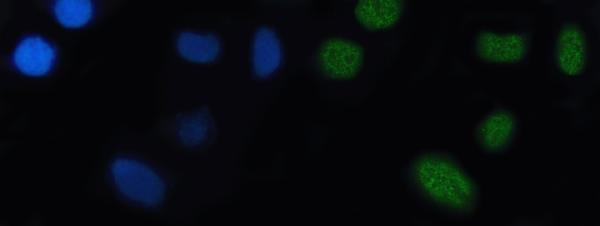
Nucleus.




PMID: 8900537 by Turbay D., et al. Molecular cloning, chromosomal mapping, and characterization of the human cardiac-specific homeobox gene hCsx.
PMID: 22560297 by Koss M., et al. Congenital asplenia in mice and humans with mutations in a Pbx/Nkx2- 5/p15 module.