This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

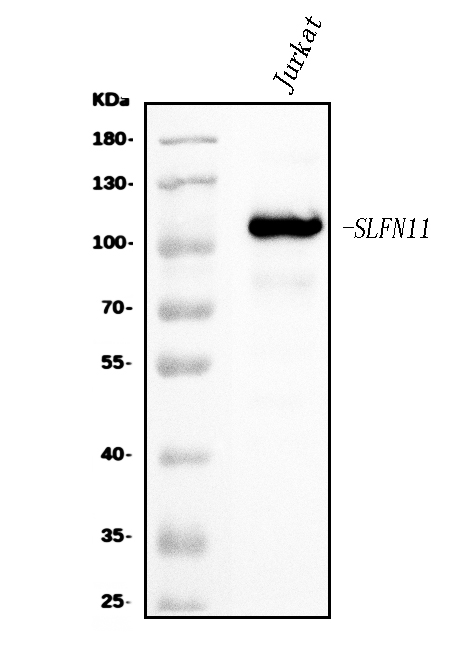
Facts about Schlafen family member 11.

Persistently blocks stressed replication forks by opening chromatin across replication initiation sites at worried replication forks, possibly leading to unwind DNA ahead of the MCM helicase and obstruct fork progression, ultimately leading to cell death (PubMed:29395061). Acts independently of ATR (PubMed:29395061).
| Human | |
|---|---|
| Gene Name: | SLFN11 |
| Uniprot: | Q7Z7L1 |
| Entrez: | 91607 |

| Belongs to: |
|---|
| Schlafen family |

FLJ34922; schlafen family member 11
Mass (kDA):
102.836 kDA

| Human | |
|---|---|
| Location: | 17q12 |
| Sequence: | 17; NC_000017.11 (35350305..35373914, complement) |
Exhibits a wider expression range in ovarian and colon adenocarcinoma than in their corresponding healthy tissues.
Nucleus. Chromosome. Recruited to stressed replication forks carrying extended RPA filaments (PubMed:29395061). Recruited to DNA damage sites via interaction with RPA1 (PubMed:26658330, PubMed:29395061).



PMID: 20956525 by Katsoulidis E., et al. Role of interferon {alpha} (IFN{alpha})-inducible Schlafen-5 in regulation of anchorage-independent growth and invasion of malignant melanoma cells.
PMID: 23000900 by Li M., et al. Codon-usage-based inhibition of HIV protein synthesis by human schlafen 11.