This website uses cookies to ensure you get the best experience on our website.
- Table of Contents

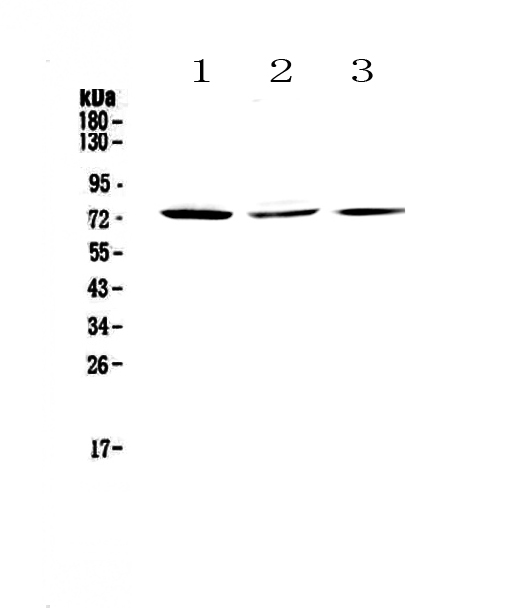
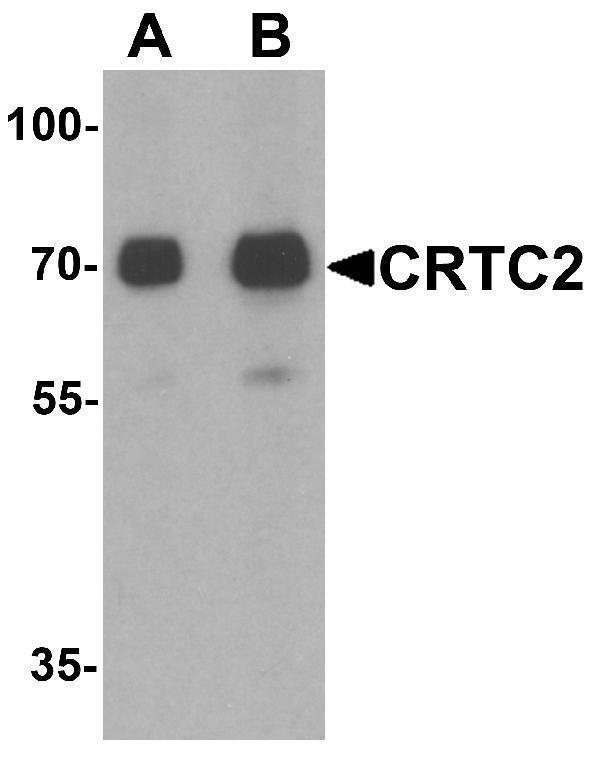
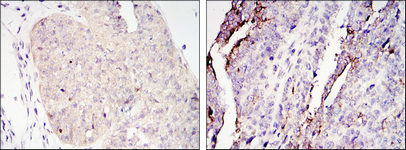
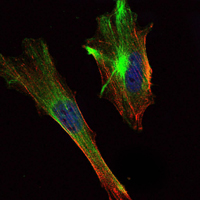
Facts about CREB-regulated transcription coactivator 2.
Enhances the interaction of CREB1 with TAF4. Regulates gluconeogenesis as a component of the LKB1/AMPK/TORC2 signaling pathway.
| Mouse | |
|---|---|
| Gene Name: | Crtc2 |
| Uniprot: | Q3U182 |
| Entrez: | 74343 |

| Belongs to: |
|---|
| TORC family |

CREB regulated transcription coactivator 2; CREB-regulated transcription coactivator 2; CRTC2; TORC2; TORC-2; TORC2transducer of regulated cAMP response element-binding protein (CREB) 2; Transducer of CREB protein 2; Transducer of regulated cAMP response element-binding protein 2
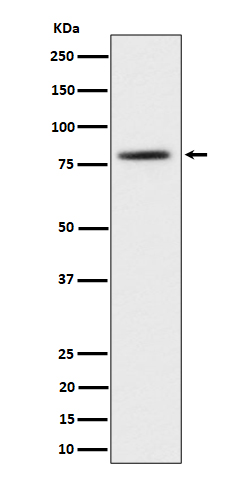
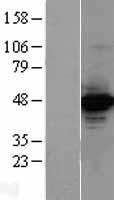
Mass (kDA):
73.216 kDA

| Mouse | |
|---|---|
| Location: | 3|3 F1 |
| Sequence: | 3; |
Expressed in the suprachiasmatic nucleus (SCN) of the brain.



PMID: 16148943 by Koo S.-H., et al. The CREB coactivator TORC2 is a key regulator of fasting glucose metabolism.
PMID: 16308421 by Shaw R.J., et al. The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin.