This website uses cookies to ensure you get the best experience on our website.
- Table of Contents
Variable and constant regions give antibodies their precision and power. Explore how each component contributes to antigen recognition, immune signaling, and the development of therapeutic antibodies used in modern medicine.
The antibody structure enables each molecule to recognize a specific target. This target is usually a unique part of a foreign substance, such as a virus, bacterium, or toxin. Once identified, the antibody helps initiate the appropriate immune response to neutralize or eliminate the threat.
Precision and action work together because of the protein's shape. Each antibody has a variable fragment that binds specifically to its target. This precise binding ensures that the immune system responds only to harmful invaders, thereby preventing it from reacting to harmless substances. After binding, antibodies can block the target’s function, mark it for destruction, or activate other parts of the immune system. These actions help the body defend itself efficiently against a wide range of threats.
Monoclonal antibodies have become one of the most widely used formats in drug development over the past two decades. In recent years, regulatory agencies have continued to approve more mAbs each year, underscoring their importance in research and clinical practice. Many of these therapies perform better after scientists adjust their variable or constant domains. Comparing the two regions, we can see why antibodies bind so accurately and why this accuracy is crucial in both research and clinical care.
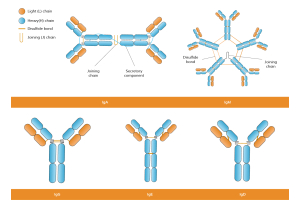
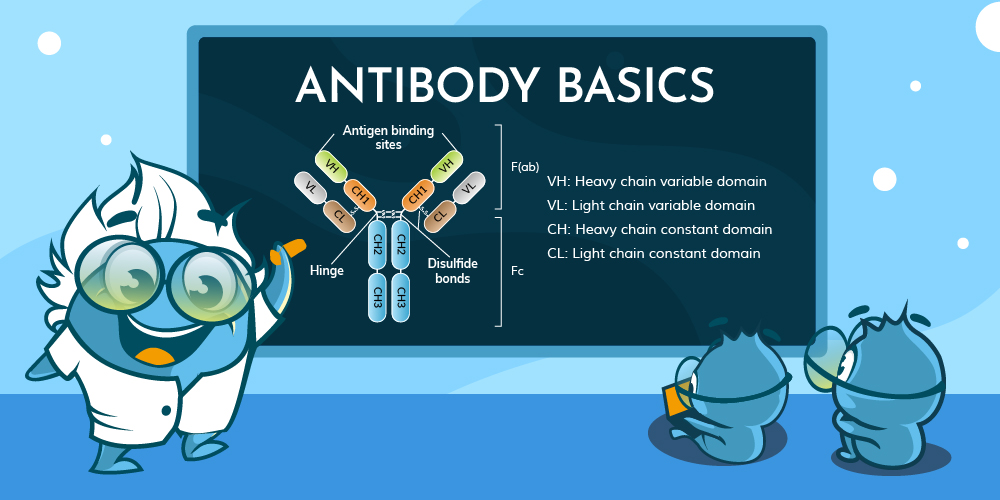
Antibodies, also known as immunoglobulins, are Y-shaped proteins built from two identical heavy and light chains. Disulfide bonds and disulfide bridges hold the chains together, giving the molecule stability and flexibility. Each chain contains a variable region at the tip and a constant region closer to the base. The variable region forms the antigen-binding site, whereas the constant region controls downstream immune activity.
Antigen recognition starts in the variable region, where sequence diversity produces unique binding surfaces.
Variable regions sit at the ends of both heavy and light chains. Within them, three complementarity-determining regions (CDRs) on each chain create the contact points for an antigen. Framework segments surround the CDRs, supporting the loops that enable binding. These hypervariable regions are critical for ensuring specificity.
When an antibody encounters its target, the six CDRs come together to form a grip on the antigen, much like a lock and key. That tight fit determines affinity, which influences how well the antibody can neutralize or flag the threat, an essential part of the antibody-antigen interaction.
Once the variable region locks onto a target, the constant region calls in reinforcements.
Constant regions extend from the hinge region of the antibody to the base of the heavy and light chains. In heavy chains, the constant segment determines the antibody isotypes, such as IgM antibodies IgG or IgA, each with distinct immune functions.
The constant region interacts with Fc fragment receptors on immune cells, activates the complement cascade, and influences serum half-life. For instance, IgG1 antibodies bind to Fcγ receptors on macrophages, guiding phagocytosis, while IgG4 has reduced effector activity, making it useful in chronic inflammatory settings.
Although neighbors within the same molecule, the two regions differ in almost every way.
| Feature | Variable Region | Constant Region |
|---|---|---|
| Primary role | Antigen recognition | Immune signalling, stability |
| Sequence variability | High | Low within an antibody class |
| Location | Tips for heavy chains and light chains | Lower arms and stem of the Y-shape |
| Flexibility | Greater to accommodate diverse targets | More rigid to maintain structural integrity |
| Engineering focus | Affinity tuning, specificity of antibody | Half-life, effector modulation |
Knowledge of the region function guides everything from laboratory assays to drug design.
Researchers often isolate variable regions to construct Fab fragments, single-chain variants, or nanobodies for use in imaging and diagnostic tests. Smaller antibody fragments retain antigen-binding activity specificity while reducing background noise.
Drug developers adjust constant regions to change how an antibody behaves in the body. For example, modifying Fc fragment domains of IgG molecules can extend half-life via neonatal Fc receptor recycling. Conversely, reducing Fc receptor binding can dampen inflammatory responses in therapeutic antibodies used to treat autoimmune diseases.
Tailoring both regions lets scientists create bispecific antibodies, antibody-drug conjugates, and chimeric antigen receptor constructs. Each relies on precise control of antigen binding and effector functional activity to improve safety and potency. Some designs also incorporate the J chain, especially in polymeric antibodies like IgM and IgA, influencing their quaternary structure and immune function.
Variable and constant regions work together, yet their tasks remain distinct. The variable portion seeks out the threat, while the constant portion signals for help. A clear understanding of both parts enables researchers to fine-tune antibodies for use in science, diagnostics, and treatment.
This includes optimizing the hinge region for structural flexibility, improving hypervariable regions for more accurate antigen recognition, and refining the amino acid sequence of polypeptide chains...
Antibody types form the foundation of adaptive immune system. In humans, IgG alone accounts for approximately 75% of circulating antibodies, while the other four classes—IgM antibodies, IgA, IgE, and IgD—fill specialized roles in blood, tissues, and secretions. Together, these Y-shaped glycoproteins create a flexible defense network that identifies, neutralizes, and clears pathogens with remarkable precision.
All antibodies share a common heterotetrametric structure consisting of heavy and lights connected by disulfide bonds in the hinge regions but variations in their constant regions define each antibody isotype. Understanding these differences is critical for selecting the right reagents for immunodiagnostic tests, designing effective vaccines, and developing therapeutic monoclonal antibodies. Each type plays a specific role in the adaptive immune response, inf...

This article summarizes comparative experimental findings and literature evidence regarding antibody validation and reproducibility in non-model organism research.


When was the last time you read the buffer section of your protocol instead of just skipping to the fun part—primary antibody incubation? Let’s be honest: for many of us, the blocking step is that quiet moment between coffee and confusion. But while often overlooked, the blocking agent you choose can make or break your experiment—and one of the most dependable names in that game is Bovine Serum Albumin, or BSA.
In this article, we take a deep dive into the science and subtlety of this humble protein—from its origins to its performance in dilution buffers, and how it stacks up against its rivals like non-fat dry milk and fish gelatin.
The story of Bovine Serum Albumin (BSA) stretches back to the 19th century, when German and Swedish chemists first separated serum proteins such as albumins and globulins from animal blood. The term “albumin” itself was already in use by the early 1800s, when French chemist Antoine Fourcroy and his contemporaries described this class of water-soluble, heat-coagulable proteins. As protein chemistry advanced, BSA was eventually isolated on its own in the late 19th to early 20th century. By the 1930s–40s, American scientist Edwin J. Cohn developed large-scale purification methods through isoelectric precipitation and plasma fractionation (the famous “Cohn fractionation”), establishing BSA as a reliable tool for experimental research. During World War II, U.S. military and NIH efforts to develop blood plasma substitutes further accelerated improvements in protein purification. While human serum albumin became the standard for clinical use, the purification of BSA played a critical role in refining protein separation technology and cemented its place in laboratory science.
Following its historical development and large-scale purification, Bovine Serum Albumin (BSA) is recognized today as a well-characterized globular protein derived from cow blood serum. As the most abundant protein in bovine plasma, BSA plays essential physiological roles in the animal, including maintaining osmotic pressure and transporting fatty acids, hormones, and other small molecules.
From a molecular perspective, BSA is approximately 66.5 kDa in size and consists of 583 amino acids arranged into a heart-shaped three-domain structure. This configuration imparts remarkable stability across a wide pH range (pH 4–9) and thermal resilience, features that make BSA particularly suitable for in vitro experimental applications. Beyond its structural robustness, BSA’s chemical inertness, high solubility, and compatibility with sensitive immunoassays underpin its ubiquitous presence on laboratory benches worldwide.
In immunoassays like ELISA, Western blot, and IHC, the blocking step prevents antibodies from binding nonspecifically to unoccupied surfaces. If ignored or done poorly, the result is often high background noise and unreliable data.
Enter BSA. Its widespread use as a blocking agent is thanks to three main features:
• Low cross-reactivity: BSA is unlikely to bind to antibodies or interfere with antigen-antibody interactions, especially in mammalian systems.
• Surface coverage: Its globular nature helps it evenly coat unbound plastic or membrane surfaces.
• Chemical compatibility: BSA tends to remain stable across a wide range of buffer systems and temperatures, and doesn’t degrade easily under typical assay conditions.
In essence, BSA acts like an invisible wallpaper—it quietly occupies all the real estate your antibodies might otherwise stick to accidentally, making sure only the intended interactions show up on your blot or plate.
Beyond blocking, BSA is frequently added to antibody dilution buffers. But its role here goes beyond background suppression. In these scenarios, BSA offers protein stabilization, antibody preservation, and reduction of denaturation risk, especially in working solutions that may be stored for extended periods or exposed to slight agitation or heat.
At concentrations around 0.1%–1%, BSA helps maintain antibody structure and function, especially for sensitive monoclonal antibodies or those used at very low concentrations. It also reduces the risk of antibody adsorption to plastic tubes or pipette tips—a subtle but significant source of signal loss in low-volume experiments.
Of course, BSA isn’t the only protein blocker on the scene. Here’s how it compares with several popular alternatives:
| Blocking Reagent | Origin | Cross-reactivity Risk | Fluorescence Compatibility | Shelf Stability | Typical Use Cases |
|---|---|---|---|---|---|
| BSA | Bovine serum | Low | High | Excellent | ELISA, WB, IHC, IF |
| Non-fat dry milk | Skim milk (casein) | Moderate (due to IgG) | Poor with HRP & phospho | Moderate | Western blot |
| Fish gelatin | Cold water fish | Low | Good | Moderate | IF, IHC (fluorescent dyes) |
| Normal goat serum | Goat plasma | Medium | Good | Fair | IHC/IF (species-matched) |
| Casein | Milk protein | High (esp. for phospho) | Poor with phospho detection | Moderate | ELISA (non-phospho) |
Key Takeaways:
• BSA is ideal for experiments requiring low background and high consistency.
• Non-fat dry milk is cheaper but riskier in sensitive detection systems.
• Fish gelatin is a strong contender in fluorescent applications.
• Serum-based blockers introduce species-specific variables and should be matched carefully.
Despite its popularity, BSA isn’t perfect for every situation.
• Biotin-based assays: BSA contains trace levels of biotin, which can interfere with avidin-biotin systems, leading to false positives or increased background noise.
• Phospho-specific antibody work: Some BSA formulations may contain minor impurities th


Buffers are a staple in nearly every molecular biology experiment—but not all buffers are created equal. When working with antibodies, ELISA, IHC, IF, or IP, the difference between PBS and PBST, or TBS and TBST, can greatly affect your results.
In this quick guide, we’ll compare four of the most common buffers used in immunoassays and help you choose the right one for your experiment.
| Name | Description | Basic Components | Common Uses | Characteristics | Recommended Use Cases |
|---|---|---|---|---|---|
| PBS | Phosphate-Buffered Saline | NaCl + K |